Contenido del artículo
V ANTIGENOS DE HISTOCOMPATIBILIDAD Y GENES DE LA RESPUESTA INMUNE
- Estructuras y antígenos del complejo principal de histocompatibilidad
- Función del CPH en la respuesta inmune
- REACCION MIXTA DE LINFOCITOS
- PROCESO Y PRESENTACION DEL ANTIGENO
- SELECCION CLONAL DE LAS CELULAS T
- PRODUCCION DE LINFOCITOS T CITOTOXICOS
- Genes de la respuesta inmune
- Complejo principal de histocompatibilidad y enfermedades
DR. CHARLES B. CARPENTER
Estructuras y antígenos del complejo principal de histocompatibilidad
Los resultados de los estudios de transplante de tumores e injertos en ratones dieron lugar a la ciencia de la inmunogenética. Si un ratón singénico proveniente de una cepa A era sometido a la aplicación de un injerto de un ratón de la cepa B, rechazaría dicho injerto. De manera similar, el ratón A produciría anticuerpos que podrían destruir a las células del ratón B (en presencia de complemento). Los antígenos de membrana reconocidos por tales anticuerpos son codificados por genes situados en loci estrechamente relacionados en el cromosoma 17, en una región conocida como H-2. A esta región cromosómica, cuyos análogos son también responsables del rechazo de injertos en otros mamíferos, se le denomina complejo principal de histocompatibilidad (CPH).
En los humanos se encuentran anticuerpos contra antígenos codificados por el CPH en el suero de multíparas y en los receptores de múltiples transfusiones sanguíneas. Estos sueros aglutinan o producen lisis de leucocitos de algunas personas, pero no de otras. En los humanos, el CPH se denomina con las siglas HLA y comprende diversos loci situados en el brazo corto del cromosoma 6 [ver figura 1].
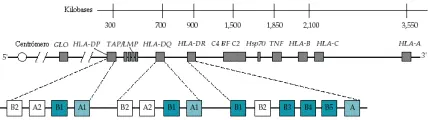
|
| Figura 1 |
| Loci del complejo principal de histocompatibilidad |
Existen dos tipos estructurales de moléculas en el CPH, moléculas de clase I y de clase II. Las moléculas de ambas clases participan en el reconocimiento de antígenos y ayudan a enfocar las defensas inmunológicas contra las invasiones de microrganismos. También participan en la comunicación entre las células durante la respuesta inmune. Las moléculas del CPH actúan mediante la unión a antígenos potenciales, que suelen ser fragmentos de péptidos que han sido procesados en las células presentadoras de antígenos. Los receptores antigénicos situados en las células T, determinados en forma clonal, reconocen y se unen a complejos peptídicos específicos del CPH, desencadenando la respuesta inmune apropiada. Los segmentos de las moléculas del CPH muestran secuencias homólogas con inmunoglobulinas, antígenos receptores de células T y moléculas de interacción de la célula T como CD4 y CD8, lo cual indica que todas estas moléculas comparten un origen común en la evolución.
ANTIGENOS DEL COMPLEJO PRINCIPAL DE HISTOCOMPATIBILIDAD
De manera tradicional, el término antígeno del CPH se ha aplicado al producto de un locus genético que muestra polimorfismo en una población de individuos. En la actualidad se ha dilucidado en forma extensa la secuencia y estructura de las moléculas que portan los antígenos del CPH, y se ha determinado que las porciones polimórficas, o antigénicas, de las moléculas del CPH son muy pequeñas. De hecho, las porciones polimórficas con frecuencia consisten en sólo una a cuatro sustituciones de aminoácidos codificadas en regiones de la secuencia hipervariable de nuclétidos del ADN. La configuración específica en una molécula del CPH causada por las sustituciones de aminoácidos determinados se conoce como epítope.
Antígenos clase I del CPH
Los antígenos clase I del CPH consisten en dos cadenas polipeptídicas unidas entre sí por enlaces no covalentes. Una cadena es pesada y glucosilada (44 kd), y determina la especificidad antigénica. La porción extracelular de esta cadena pesada clase I se divide en tres dominios designados alfa1, alfa2 y alfa3. La otra cadena es una proteína pequeña (11.5 kd) conocida como ß2-microglobulina [ver figura 2]. En los ratones, los antígenos de clase I son codificados por genes de loci denominados H-2K y H-2D, así como en otros loci.
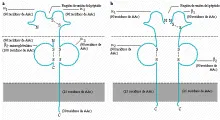
|
| Figura 2 |
| Moléculas del complejo principal de histocompatibilidad |
En los humanos las cadenas pesadas de clase I son los productos de los genes de tres loci del CPH denominados HLA-A, HLA-B y HLA-C. Existen muchos alelos para cada locus y, por consiguiente existe considerable polimorfismo [ver tabla 1]. La ß2-microglobulina se codifica en un gen situado en el cromosoma 15. Tanto la ß2-microglobulina como el dominio alfa3 de la cadena glucosilada pesada de los antígenos de clase I del CPH muestran una semejanza estructural considerable en la región constante de la cadena pesada de la IgG (CH3).
|
||||||||||||
* Principales genes espresados de los que se
conocen sus secuencias. |
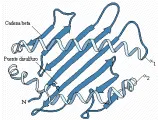
La moléculas de clase I ya se han cristalizado y su estructura se ha determinado mediante difracción de rayos-X a una resolución de 3.5 A [ver figura 3].1 Se encontró que dos de los dominios de las cadenas pesadas, alfa1 y alfa2, se localizan en la porción distal de la membrana de la cadena pesada y forman un surco a lo largo de la superficie superior de la molécula. Los lados del surco están compuestos de hélices-alfa de los dominios alfa1 y alfa2, y la base se compone de ocho placas antiparalelas con plegamiento ß de estos dominios. Las regiones hipervariables (antigénicas) se encuentran sobre todo a lo largo de los lados del surco, pero existe también cierta variabilidad en la región de la placa ß. El resto de la molécula muestra mínima variabilidad en relación con otras moléculas del mismo locus HLA. En los cristales estudiados el surco que se encuentra fuera de la membrana celular y tiene una longitud aproximada de 25 A por 10 A de ancho, contiene una molécula no identificada que al parecer representa antígeno procesado (i.e., fragmentos peptídicos). Cuando se separan estos péptidos de las moléculas de clase I purificadas, y se determina su secuencia, muestran patrones de aminoácidos, denominados motivos, que se fijan a grupos especiales de moléculas de clase I del HLA.2 Estos datos ayudan a confirmar la hipótesis de que las moléculas del CPH unen y presentan antígenos procesados a las células T que responden, y que el receptor de células T (RCT) reconoce a los antígenos extraños como un péptido en el contexto de autoantígenos; esto es, que se une a una superficie compuesta por el CPH y por el péptido unido.
|
|
|

Antígenos de clase II del CPH
Algunos anticuerpos obtenidos por inmunización con células histocompatibles reaccionan con una variedad limitada de células, sobre todo linfocitos B, monocitos, células dendríticas y linfocitos T activados. En condiciones normales se ha demostrado que solo estas células poseen antígenos de clase II del CPH. Sin embargo, al igual que con los antígenos de clase I, los estados inflamatorios hacen que muchos otros tejidos expresen antígenos de clase II.
Los antígenos de clase II consisten en dos polipéptidos glucosilados introducidos en la membrana y denominados alfa (34 kd) y ß (28 kd); estos polipéptidos se encuentran unidos en forma no covalente [ver figura 2]. La porción extracelular de la cadena alfa se divide en dos dominios denominados alfa1 y alfa2; la porción extracelular de la cadena ß también se divide en dos dominios ß1 y ß2. En los ratones los antígenos de clase II del CPH son codificados por un gen en la región referida con la letra I. En los humanos, los antígenos de clase II son codificados por genes de la región HLA-D, la cual se divide cuando menos en tres subregiones: HLA-DP, HLA-DQ y HLA-DR [ver figura 1].
Los estudios cristalográficos indican que las moléculas de clase II del CPH poseen una estructura similar a las moléculas de clase I del CPH, formando un surco con los dominios alfa1 y ß1, en el que las placas ß forman la base y las hélices alfa forman los lados [ver figura 3].3 Al igual que las moléculas de clase I del CPH, las regiones hipervariables (antigénicas) se localizan sobre todo a lo largo del surco, lo que indica la existencia de una base molecular para el correconocimiento por el RCT del antígeno extraño y del CPH propio.
Los ratones poseen genes para dos tipos de moléculas de clase II, uno en el locus I-A (que contiene genes para las subunidades Aalfa y Aß) y otro en el locus I-E (que codifica las subunidades Ealfa, Eß y Eß2). La subdivisión de la región clase II del ratón ha sido establecida por estudios realizados en ratones congénicos, apareados de manera que sean genéticamente idénticos excepto en porciones del complejo principal de histocompatibilidad. Es posible aparear ratones con alelos idénticos K y D (clase I) pero con diferencias en las regiones II. Mediante inmunización cruzada se obtienen anticuerpos contra diversos antígenos codificados en el loci A y E de la región de clase II.
En los humanos pueden identificarse antígenos similares utilizando sueros de mujeres multíparas que reaccionan sobre todo con linfocitos B. Los sueros se exponen primero a plaquetas obtenidas de un reservorio de muchos individuos, ya que las plaquetas contienen antígenos de clase I pero no de clase II, por lo que adsorberán los anticuerpos contra los antígenos de clase I, dejando a los anticuerpos contra los antígenos de clase II en el suero. En la actualidad, la nomenclatura de los genes de la región HLA-D se basa en el conocimiento de la bioquímica de los antígenos expresados y en la creciente base de datos de la secuencia de nucleótidos del ADN. Por ejemplo, el gen que codifica la cadena alfa del HLA-DR se denomina DRA. De manera similar, los genes ligados estrechamente que codifican las cadenas ß se han denominado DRB1 (que codifica las cadenas ß para DR1 a DR18), DRB3 (que codifica las cadenas ß para DRw52), DRB4 (que codifica las cadenas ß para DRw53) y DRB5 (que codifica una cadena ß para Dr51). Debido a que DRB2 no expresa productos proteicos, se le denomina seudogén. Cada una de las cadenas ß del HLA-DR se relaciona con la cadena alfa común no polimórfica para formar moléculas funcionales de clase II del HLA-DR. En vista de que las cadenas alfa del DRA son siempre las mismas, la diferencia en los alelos del HLA-DR se explica por las variaciones en los genes que codifican las cadenas ß del DR. La subregión HLA-DQ contiene los genes DQA1, DQB1, DQA2 y DQB2. Al parecer DQA2 y DQB2 son seudogenes, mientras que los productos de DQA1 y DQB1, que son las cadenas alfa y ß del HLA-DQ, son polimórficas. La organización del gen HLA-DP es semejante a la del HLA-DQ [ver figura 1].
Análisis de la secuencia y nomenclatura del HLA
La mayoría de los alelos del sistema HLA pueden identificarse por medio del análisis de fragmentos de restricción de longitud polimórfica (FRLP), una técnica que consiste en la electroforesis de los fragmentos del ADN después de que éste se ha expuesto a diversas endonucleasas de restricción.4 La técnica de reacción en cadena de la polimerasa proporciona un método más preciso y adecuado para tipificar a los alelos del HLA que el análisis de los FRLP. En este procedimiento los oligonucleótidos principales específicos para ciertos locus (v.gr., DRB1, DQA1) son utilizados para estimular de manera selectiva la amplificación de los segmentos del ADN que incluyen las regiones variables que codifican para alelos específicos. Enseguida el ADN amplificado es combinado en las sondas de oligonucleótidos para detectar la presencia de una secuencia específica.5 El método de RCP tiene la ventaja de requerir un número muy pequeño de células y, con modificaciones adicionales, debe proporcionar resultados tan rápidos como las pruebas serológicas ordinarias.
En la actualidad la información sobre las secuencias del ADN es tan amplia que los alelos del HLA son definidos de manera oficial por sus secuencias, más que por sus patrones serológicos. Se están empezando a definir las bases estructurales para la especificidad de la alorreactividad con los nuevos datos acumulados. Algunos alelos nuevos, los cuales todavía no han sido definidos serológicamente, se han identificado por análisis de secuencia [ver tabla 1]. Las diferencias de aminoácidos puede presentarse en diversos sitios sobre la hélice alfa o las placas ß [ver figura 3]. Aunque existen tres o cuatro regiones en la molécula de HLA que tienen mayor probabilidad de variar (las denominadas regiones hipervariables), solo una o dos sustituciones de aminoácidos son suficientes para producir una diferencia serológica. La formación de secuencias también proporciona cierto grado de información acerca de los polimorfismos del HLA-Dw, el cual se ha definido de manera clásica solo por su reactividad en la reacción mixta de linfocitos en cultivos de tejidos (ver adelante). En la nueva nomenclatura el acuerdo es anotar primero el locus, seguido por un asterisco (*) y después por un número de cuatro dígitos. Los primeros dos dígitos son el número del alelo, y los dos últimos dígitos indican una secuencia única para las variantes. Por ejemplo, las cuatro variantes más comunes del HLA-A2 ahora se designan como A*0201, A*0202, A*0203 y A*0204. Solamente las secuencias únicas se ajustan a la nueva nomenclatua. Otro ejemplo es el DR2, que es el antígeno predecesor de los fragmentos serológicos DRw15 y DRw16. Ahora se sabe que existen cuatro cadenas de DRB1 para DR2. En la nomenclatura, basada en la secuencia de nucléotidos, el DR2 se designa como DRB1*1501, DRB1*1502, DRB1*1601, o DRB1*1602. Los fragmentos 01 y 02 representan a las cuatro especificaciones conocidas previamente como HLA-Dw: DW2, Dw12, Dw21 y Dw22, respectivamente. Hay muchos ejemplos de correlación celular (i.e., HLA-Dw) y serológica con las secuencias HLA-A, HLA-B, HLA-C, DRB1, DRB3, DRB4, DRB5, DQA1, DQB2 y DPB1, pero no todas las secuencias pueden definirse por técnicas de tipificación serológica.6
Frecuencia de los diferentes alelos del HLA
Dos términos, haplotipo y desequilibrio de unón, describen asociaciones importantes entre los genes del CPH. Haplotipo se refiere al conjunto de genes de cualquier cromosoma. Cada individuo posee dos haplotipos del complejo principal de histocompatibilidad, uno de cada progenitor. Cada haplotipo posee una serie de antígenos en particular, determinada por los loci HLA-A, HLA-B, HLA-C, HLA-DR y por otros más.
El segundo término, desequilibrio de unión, se refiere a la observación de que algunos antígenos del HLA coinciden con un solo haplotipo con mucha mayor frecuencia que la esperada. Si se distribuyen genes aislados en forma independiente a través de la población, la frecuencia con la que se combinaría cualquier par de antígenos relacionados, codificados en locus diferentes dentro de un haplotipo, sería simplemente el producto de su frecuencia en la población. Sin embargo, el antígeno HLA-A1 y el antígeno HLA-B8 se asocian con una frecuencia de seis a 21 veces mayor de lo esperado por la frecuencia de sus genes. Este desequilibrio de unión tal vez ocurra porque no ha transcurrido suficiente tiempo en la evolución para que los genes que gobiernan los antígenos se distribuyan en forma normal, o porque tal relación resulte en una ventaja para el individuo. La recombinación, o entrecruzamiento, tiene lugar durante la meiosis y se presenta en cerca del 1.5 porciento de las veces entre los locus de clase I y de clase II [ver figura 1]. Después de muchas generaciones, estas recombinaciones llegan a un equilibrio entre los alelos asociados en una población, a menos que la influencia selectiva favorezca la supervivencia de ciertos haplotipos. Un ejemplo hipotético de esta selección lo representa la supervivencia de individuos relacionados con haplotipos HLA que le confieren resistencia a padecimientos epidémicos, como la viruela o la peste. Las diferencias raciales se reflejan en las variaciones observadas en la frecuencia de ciertos antígenos y haplotipos HLA. Pocos, aunque a menudo notables, son los ejemplos de estas diferencias que se han observado en varios grupos étnicos.
Función del CPH en la respuesta inmune
REACCION MIXTA DE LINFOCITOS
Cuando se cultivan los linfocitos de un individuo junto con los de otro, las células son estimuladas para dividirse. Esta división, que puede medirse vigilando la velocidad de captación de timidina marcada con 3H en el ADN celular, se denomina reacción mixta de linfocitos (RML). Impidiendo la división de un grupo de estas células mediante tratamiento con mitomicina o irradiación, es posible estudiar a los antígenos de membrana de las células tratadas que estimulan esta respuesta proliferativa. En los ratones, las moléculas de clase II codificadas en la subregión I-A e I-E portan los antígenos de la RML. En los humanos, después de que se definieron en forma serológica los loci de clase II HLA-DR, HLA-DP y HLA-DQ, se encontró que los determinantes antigénicos HLA-DR parece ser los que más influyen en la evocación de una RML primaria. Los antígenos HLA-DQ desempeñan algún papel, mientras que los antígenos HLA-DP no parecen participar en la RML primaria. Sin embargo, los linfocitos de respuesta que se han preparado por exposición previa a antígenos HLA-DR, HLA-DQ o HLA-DP proliferan en forma vigorosa cuando vuelven a exponerse al mismo antígeno en una RML secundaria. Por lo tanto, las células T se comportan como si ya fueran inmunes a otro HLA-DR individual, lo que sugiere que la presentación de péptidos por moléculas DR tiene gran posibilidad de causar inmunidad.
PROCESO Y PRESENTACION DEL ANTIGENO
La transformación de proteínas en fragmentos peptídicos es una parte importante del proceso por medio del cual los antígenos son presentados a las células T y a otras células de la respuesta inmune. Las moléculas del CPH de la superficie celular se comportan como si tuvieran péptidos ya fijados. El desplazamiento por péptidos adheridos puede observarse en las moléculas de clase II, pero no en las moléculas de clase I. Por lo tanto, parece ser que las proteínas son primero degradadas en el interior, y los fragmentos peptídicos se unen a moléculas de clase I y de clase II del CPH dentro de la célula. Las moléculas de clase I se expresan en prácticamente todos los tejidos. Las células infectadas por virus son reconocida principalmente por células T con restricción de clase I, por lo general los que tienen función citotóxica. Por el contrario, las células T dirigidas contra moléculas de clase II interactúan con células presentadoras de antígeno del sistema inmune (i.e., células B, macrófagos, células dendríticas o células de Langerghans) que se ocupan especialmente de la defensa contra agentes infecciosos externos. Debido a que las células que poseen antígenos de clase II también expresan antígenos clase I, pueden actuar como células presentadoras de antígeno para proteínas tanto endógenas como exógenas.
Los antígenos exógenos y endógenos alcanzan la superficie celular por diferentes vías.7 Las proteínas exógenas son captadas por los endosomas o los lisosomas, donde son catabolizadas. Por lo general, estos péptidos de las proteínas exógenas se unen a las moléculas de clase II del CPH, y enseguida los complejos péptido-molécula clase II del CPH son llevados a la superficie para su presentación a las células T. Los péptidos de las proteínas endógenas (v.gr., proteínas secretadas o productos de infecciones virales) parecen formar complejos en el retículo endoplásmico con las moléculas clase I del CPH. Genes que también se localizan en la región del CPH, llamados LMP, codifican proteínas que son las responsables de fragmentar las proteínas en pequeños péptidos (con longitud de ocho a 10 aminoácidos); los genes TAP, muy relacionados, codifican chaperones que transportan los péptidos a través de las membranas intracelulares [ver figura 1].8 El sistema lleva péptidos de origen intracitoplásmico a las moléculas de clase I recién formadas. Como ya se mencionó, ciertas secuencias de los péptidos son características de los péptidos obtenidos de moléculas purificadas de un determinado alelo del CPH. Estos datos indican que las diferencias en la secuencia alélica en los márgenes del surco de unión del péptido determinarán qué peptido se une. Los péptidos unidos a moléculas clase I suelen tener nueve aminoácidos, con residuos en sitios especiales que tienen una carga similar o hidrofobicidad (v.gr., en las posiciones 1, 3 y 9)2,9 para diferentes grupos de alelos HLA. Además, un número de péptidos sintéticos que representan porciones inmunogénicas de agentes infecciosos u otras proteínas extrañas también se alinean en motivos comunes semejantes. Los péptidos obtenidos de moléculas purificadas de clase II varían en longitud hasta 25 residuos y tienen una longitud mínima de 13 a 14 aminoácidos.10 Los motivos para el DR1 tienen un residuo con carga positiva en la posición 1, un donador de enlace hidrógeno en la 6 y un residuo hidrofóbico en la 10. En el futuro podrá ser posible predecir la afinidad de una secuencia de unión determinada para cada molécula del CPH.
SELECCION CLONAL DE LAS CELULAS T
El reconocimiento simultáneo del CPH propio y de los fragmentos de un antígeno (X) unido en el surco de las moléculas de clase I o II parece requerir que la superficie de unión del receptor de la célula T y la superficie de unión formada por el CPH más péptido se fijen en múltiples puntos. Cada clona de células T es específica para el CPH propio más X y de manera general no tiene suficiente afinidad para unir al CPH o al X en forma aislada. Hay numerosos datos de que el desarrollo de la variedad de células T en el timo inicia en el periodo fetal y continúa así hasta la vida adulta, cuando nuevas células precursoras de la médula ósea son preparadas en el timo. En este proceso muchas clonas potenciales son destruidas y otras son seleccionadas para madurar. Posteriormente las clonas de células T seleccionadas se desplazan para poblar al resto del organismo.11
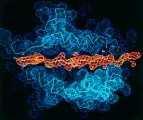
El CPH del huésped desempeña una función importante en la selección: las clonas de células T que son muy autorreactivas a moléculas de CPH propios son eliminadas, sobreviviendo aquellas clonas con débil afinidad al CPH propio. En vista de que los supervivientes tienen una gran variedad de distribuciones de los receptores de células T, persiste la variedad necesaria de clonas de células T que pueden reconocer al CPH propio más el péptido antigénico. La cirstalización exitosa de un complejo que consiste en un RCT humano, su péptido viral y la molécula HLA-A2 que se une a éste, ha revelado la configuración y extensión de la superficie de unión entre el RCT y el CPH más el péptido.12 El eje del RCT es diagonal al de las hélices del CPH, y cubre una gran porción de ambas helices alfa y al péptido localizado entre ellas [ver figura 4]. Aunque los muchos polimorfismos del CPH aumentan la probabilidad de que un fragmento peptídico determinado pueda ser unido y reconocido por las células T, un individuo determinado tiene una variedad relativamente pequeña de sitios de unión de CPH en comparación con las múltiples posibilidades de combinación en el complejo del gen del RCT. Sin embargo, la trasmisión hereditaria de múltiples locus HLA de los dos padres aumenta la posibilidad de reconocer un número mayor de complejos CPH propios más péptidos diferentes y, por lo tanto, aumenta la probabilidad de que un individuo sobreviva a una infección determinada.
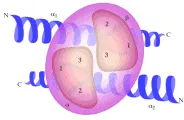
|
| Figura 4 |
| Receptor de la célula T |
La alorrespuesta, que es la respuesta inmune desarrollada contra células de otro individuo, es un caso especial. Excepto por activación directa de grupos de células T con superantígenos bacterianos (v.gr., exotoxinas estafilocóccicas) la proliferación in vitro de células T en las RML es la respuesta antígeno-específica más intensa que se conoce en vista de que no requiere preparación necesaria para inducir la proliferación contra los antígenos microbianos. Por supuesto el transplante es un modelo clínico y experimental que no habría surgido per se durante la evolución; solo el embarazo tiene el potencial para exponer las células de un individuo a otro de diferente haplotipo HLA. La alobarrera pudo haber hecho difícil o imposible el embarazo durante la evolución, excepto por la presencia de varios mecanismos mal definidos a nivel placentario que protegen al feto del rechazo. La existencia de estos mecanismos sugiere que la necesidad de los polimorfismos del CPH es muy importante y que se requiere protección especial en la interfase madre-feto.
Se sabe que las células T alorreactivas perciben en forma indirecta a los halopéptidos-CPH presentados sobre las moléculas del CPH propio o reconocen de manera directa a las moléculas del alo-CPH intactas que contienen a los péptidos propios.13 En vista de que varios péptidos derivados de proteínas endógenas probablemente ocupan los sitios de unión del CPH todo el tiempo, estos péptidos propios no necesitan ser polimórficos o únicos para un individuo. En la actualidad se investiga el significado funcional de las vías tanto indirectas como directas en el trasplante, y se ha demostrado en forma experimental que la inmunización con alopéptidos solos puede acelerar el rechazo de injerto,14 mientras que la administración de estos péptidos por vía oral o intratímica aumenta la tolerancia para los autoantígenos.15
PRODUCCION DE LINFOCITOS T CITOTOXICOS
Para que una RML conduzca a la producción de linfocitos T citotóxicos se requieren dos tipos de linfocitos T que respondan. El proceso comienza con una célula estimuladora (linfocito B, célula dendrítica o monocito) que posee moléculas de clase I y de clase II del CPH en su superficie. La molécula de clase II estimula ciertos subtipos de linfocitos T para que proliferen y se transformen en células T cooperadoras; este subtipo de linfocitos T está marcado por el antígeno CD4. Las moléculas de clase I sensibilizan a un segundo subtipo de linfocitos T, que se transforman en células citotóxicas si son estimuladas por los linfocitos T cooperadores en proliferación. Una de estas señales estimuladoras es modulada por la linfocina interleucina-2. Este segundo subgrupo está marcado por el antígeno CD8. Las células T citotóxicas que se desarrollan contra células que difieren sólo en sus antígenos de clase II son portadoras del marcador CD4. Los dos estímulos, aquel que induce la proliferación de linfocitos T cooperadores y el que sensibiliza a los linfocitos para que se vuelvan citotóxicos, pueden ser transmitidos por células diferentes [ver tabla 2]. Se supone que este tipo de interacción celular es un reflejo de los eventos in vivo que conducen al rechazo de injertos por los linfocitos T citotóxicos e ilustra la necesidad de que el donador y el receptor compartan los antígenos celulares de clase I y de clase II.
|
||||||||||||||||||||||||||
* Las células estimuladoras que inducen proliferación
de las células T respondedoras de la reacción mixta de cultivo
de linfocitos también sirven como blancos de las células
citotóxicas que se desarrollan a partir de la población de
respuesta (medidas por ensayo de linfolisis mediada por células).
|
||||||||||||||||||||||||||
Antes se consideraba que las células T CD4+ representaban tan solo linfocitos cooperadores y que las células T CD8+ eran linfocitos citotóxicos o supresores, pero estas divisiones funcionales no parecen ser tan claras. Se están realizando estudios moleculares que indican que la molécula de superficie CD4 se relaciona estrechamente con el receptor de células T y guía la interacción entre las células T y las células presentadoras de antígenos, al unirse a una región no polimórfica de las moléculas de clase II del CPH. De manera similar, la molécula CD8 se relaciona en forma estrecha con las moléculas de clase I del CPH sobre las células presentadoras de antígenos. Las moléculas CD4 y CD8 también aumenten la potencia con la que el complejo RCT se une al complejo entre el antígeno y el CPH. Además, estas moléculas de superficie pueden participar en la activación de la célula T adherente. Al considerar las funciones supresoras, la evidencia indica que las células T CD4+ inducen a las células T CD8+ supresoras, pero la relación entre las moléculas de superficie CD4 y CD8 con las moléculas del CPH todavía no se ha aclarado.
Genes de la respuesta inmune
Como ya se mencionó, muchas lineas de evidencia indican que las moléculas de clase II del CPH son los productos expresados de los genes de la respuesta inmune (Ir); en otras palabras, la respuesta al parecer es una función directa de la presentación del antígeno. Si un fragmento de antígeno no se une a la molécula de clase II, el sistema inmune del individuo no es capaz de reconocerlo. Algunas enfermedades de los animales, incluyendo formas de leucemia inducidas por virus, tumores mamarios y coriomeningitis linfocítica, se han relacionado con los genes de clase II del CPH. Sin embargo, no se ha demostrado la capacidad de antígenos específicos del HLA para conferir susceptibilidad a agentes infecciosos importantes desde el punto de vista clínico. Es posible que la evolución haya causado la selección de los alelos del CPH que son capaces de unirse cuando menos a algunas porciones de las moléculas antigénicas de los agentes infecciosos. Además, la duplicación de los genes de clase II con la expresión de los grupos de moléculas HLA-DR, HLA-DQ y HLA-DP aumenta la probabilidad de que una respuesta sea iniciada en un caso determinado. En especial, los polimorfismos en las cadenas tanto alfa como beta del HLA-DQ y HLA-DP proporcionan variación considerable en las configuraciones de unión, en especial cuando los dímeros alfa/beta están compuestos de cadenas heredadas de ambos padres; por ejemplo alfamadre/betapadre puede proporcionar una molécula de unión a un péptido no presente en ninguno de los dos padres. También existen muchas influencias no relacionadas al CPH en la respuesta inmune, y ninguna de éstas ha sido bien caracterizada aún desde el punto de vista clínico.
Los estudios en humanos han sugerido también la capacidad del CPH para suprimir las respuestas inmunológicas a agentes ambientales, como infección estreptocócica, esquistosomiasis y lepra, así como antígenos del polen del cedro y de la vacuna de la hepatitis B. Por lo tanto, se ha postulado la participación de genes inmunosupresores (Is). Como en el caso de los genes Ir, al parecer está implicada la región de los antígenos de clase II del CPH. Por ejemplo, la respuesta in vitro de la IgE al antígeno del polen del cedro es suprimida por células T de individuos portadores de HLA-DQ3,16 pero los mecanismos de esta supresión mediada por células T no está bien definida.
GENES DE LOS FACTORES DEL COMPLEMENTO
Varias proteínas del complemento son codificadas por genes ligados al CPH. Estas proteínas incluyen C2 y el factor B (FB) que están muy relacionados e incluso son similares desde el punto de vista estructural, lo que sugiere duplicación de genes. Además, dos loci que codifican submoléculas de C4 (C4A y C4B) están muy ligados a C2 y FB. La deficiencia de C2 asociada con lupus eritematoso generalizado se relaciona con el haplotipo Aw25-B18. De hecho, se han descrito haplotipos extendidos en los que los mismos tipos de antígenos HLA-B, HLA-DR, HLA-DQ y complemento se encuentran en personas aparentemente no relacionadas, que padecen la misma enfermedad. Estas circunstancias podrían resultar de una mutación que ocurrió en un antepasado común. En forma alternativa, puede haber presión selectiva para mantener en estrecha proximidad genes que producen proteínas que actúan en conjunto.
FUNCIONES NO INMUNOLOGICAS DE LOS GENES DEL CPH
Los genes del CPH al parecer también tienen importancia en diversas reacciones intercelulares que no son propiamente inmunológicas. En 1976 se dio a conocer un estudio en el cual se demostró que un ratón macho que se apareaba con dos hembras congénicas H-2 en etapa de celo prefería aparearse con hembras que tuvieran el tipo H-2 diferente al propio; estudios ulteriores mostraron que el macho hacía esta distinción por medio del olfato. La ventaja más aparente en este ejemplo de atracción entre polos opuestos es que la heterocigocidad de los genes de la región H-2, sobre todo de los genes Ir garantiza un mayor número de defensas inmunológicas para el producto híbrido de dichos apareamientos. Sin embargo, no existen evidencias de que los humanos pueden percibir los antígenos HLA. Aunque los antígenos de superficie controlados por el CPH pueden tener también un papel importante en la organogénesis y diferenciación, existen evidencias contrarias para esta hipótesis provenientes de estudios de ratones que no tienen expresión del CPH en la superficie de las células.
Complejo principal de histocompatibilidad y enfermedades
ENFERMEDADES RELACIONADAS CON EL HLA
Se han relacionado muchas enfermedades con ciertos antígenos específicos del CPH [ver tabla 3].17 Estas asociaciones per se muestran solo que las moléculas del CPH, o algunos otros genes estrechamente relacionados con la región HLA, tienen una influencia sobre el inicio o expresión de la enfermedad. Por ejemplo, un riesgo relativo de 5 significa solo que existe un incremento de cinco veces en la posibilidad de la enfermedad en un individuo con un antígeno HLA particular, comparado con alquien que no tiene ese antígeno. Una explicación para estas asociaciones es que la enfermedad en cuestión está relacionada con una deficiencia en la respuesta inmune a un organismo causal en particular. Sin embargo, existe evidencia creciente de que las enfermedades órgano específicas asociadas al HLA, como la diabetes mellitus insulinodependiente (DMID), la enfermedad de Graves, las glomerulonefritis, la enfermedad celiaca, la espondilitis anquilosante y la artritis reumatoide, tienen un componente de autoinmunidad importante.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
* Riesgo relativo = (% pacientes con el antígeno) (% controles sin el antígeno) |
En los modelos animales en los que se han realizado estudios apropiados, se ha demostrado que los estados autoinmunes dependen de cinco a 15 genes segregados en forma aleatoria, uno de los cuáles se encuentra en el CPH. La etiología poligénica de la autoinmunidad humana es muy probable, pero los componentes del HLA pueden ser blancos adecuados para intervención, en especial en las enfermedades en las que la presentación del HLA de un autopéptido inmunogénico es el evento clave.18 Además, con el desarrollo de inflamación, la expresión de novo de las moléculas del HLA de clase II en las células de los tejidos puede proporcionar el estímulo inmunológico para la perpetuación del proceso autoinmune. Por ejemplo, los pacientes con tiroiditis tienen expresión aberrante del HLA-DR en las células tiroideas, lo que proporciona un posible mecanismo por el que el antígeno tiroideo podría presentarse a las células T.
Existen progresos en la definición de qué enfermedades pueden relacionarse en forma directa con la presentación de los péptidos inmunogénicos. El análisis de las secuencias de los genes que codifican las moléculas de clase II del CPH de los pacientes con DMID sugiere que la herencia de los alelos específicos del HLA es importante para determinar la susceptibilidad a esta enfermedad, que puede incluir una respuesta autoinmune mediada por células T a los antigenos de los islotes pancreáticos. La resistencia a la DMID se asocia en forma estrecha con la presencia de aspartato en la posición 57 de la cadena beta del HLA-DQB.19 En los individuos con el haplotipo HLA-DR2, por ejemplo, el riesgo relativo para DMID cae a 0.2 [ver tabla 3]. El HLA-DR2 está en desequilibrio de unión con los alelos beta HLA-DQB, como el DQB1*0602, que codifica aspartato en la posición 57. Por el contrario, cuando no existe aspartato en la posición 57, en especial en individuos con el haplotipo HLA-DR3 o HLA-DR4, existe un mayor riesgo de DMID. El residuo aminoácido 57 en la cadena beta del HLA-DQB puede colocarse en un estremo del surco; el aspartato en esta posición puede influir en la unión de un péptido a esta molécula de clase II, causando reducción de las respuestas de las células T cooperadoras o activación de las respuestas de las células T supresoras a los antígenos de los islotes pancreáticos. Muchos estudios entre grupos étnicos han demostrado que la mayor susceptibilidad a la DMID se relaciona con el HLA-DQ. El heterodímero DQA/DQB DQA1*0301/DQB1*0201 se asocia con un mayor riesgo. Lo interesante aquí es que este herodímero es poco común, excepto en indiviudos que han heredado el gen DQA de un padre y DQB del otro. Mientras que los DQA1*0301 y DQB1*0201, por lo general encontrados en los haplotipos DR4 y DR3, respectivamente, aumentan en forma independiente el riesgo de DMID, juntos son responsables del mayor riesgo de la enfermedad. Como se mencionó antes, con la molécula HLA-DQ ocurre la formación de un heterodímero a partir del producto de genes heredados de ambos padres. La hipótesis consiste en que este sitio de unión para el péptido será más eficaz para presentar el autoantígeno de los islotes pancreáticos. La definición de los motivos de unión en este sitio puede proporcionar la clave sobre cuál es el antígeno. Existen otros efectos adicionales e independientes del HLA-DR, en especial de los alelos DR4, algunos de los cuales aumentan y otros suprimen el riesgo de DMID. Las diferencias de aminoácidos en las regiones hipervariables de las moléculas clase II del CPH también se han asociado con trastornos autoinmunes como pénfigo vulgar y artritis reumatoide.
La asociación de la narcolepsia con el antígeno HLA-DR2 (DRB1*1501) es mayor del 90, pero la asociación es mayor con el HLA-DQA1*0102/DQB1*0602.19 El efecto del sistema HLA es dominante, no recesivo, y no existe indicación de un defecto inmunológico en individuos afectados. Se ha postulado una alteración en un neurotransmisor o en su receptor, pero se desconoce la relación con los genes de la región HLA-D.
Entre el 80 y el 90 porciento de los casos de enfermedad celiaca se asocian con el HLADQA1*0501/DQB1*0201.20 El surco de unión de esta molécula se une a un péptido de la proteína del trigo gliadina, factor potencializador, si no etiológico, en esta enfermedad.
Aunque una molécula HLA puede determinar la especificidad a un autoantígeno en particular, es posible que los genes que controlan otros factores (v.gr., la producción de receptores de antígenos, subgrupos específicos de células reguladoras, o moléculas cooperadoras y supresoras) sean responsables de una tendencia general a una respuesta inmunológica anormal. Estudios adicionales del papel del sistema HLA en la inmunidad podrán revelar los mecanismos de la enfermedad autoinmune que no se conocen aún.
ENFERMEDADES LIGADAS AL SISTEMA HLA
Unas cuantas enfermedades están claramente ligadas a haplotipos específicos del sistema HLA en familias, teniendo en ocasiones alelos comunes en la población general y heredándose como defectos autosómico recesivos. Un ejemplo es la hiperplasia suprarrenal congénita, un defecto autosómico recesivo de la enzima 21-hidroxilasa que provoca una síntesis deficiente de cortisol y producción excesiva de hormonas androgénicas. Esta condición puede asociarse con el haplotipo HLA-B47-BFF-DR7. De manera similar, la hemocromatosis idiopática es una enfermedad autosómica recesiva caracterizada por absorción excesiva de hierro en la dieta y se relaciona con el haplotipo A3-B14. La enfermedad de Hodgkin en familias también se asocia con un patrón recesivo ligado al HLA. Debido a que la asociación con alelos específicos es débil [ver tabla 3], puede ser que exista una mutación común que ocurre en varios fundadores.
Figura 1 Laura Brown.
Figuras 2 y 3A Tom Moore.
Figura 4 Dimitry Schidlovsky.
Bibliografía
-
Bjorkman PJ, Saper MA, Samraoui B, et al: Structure
of the human class I histocompatibility antigen, HLA-A2. Nature 329:506,
1987 [PMID
3309677]
-
Sidney J, Grey HM, Kubo RT, et al: Practical, biochemical
and evolutionary implications of the discovery of HLA class I supermotifs.
Immunol Today 17:261, 1996 [PMID
8962628]
-
Brown JH, Jardetzky TS, Gorga JC, et al: Three-dimensional
structure of the human class II histocompatibility antigen HLA-DR1. Nature
364:33, 1993 [PMID
8316295]
-
Mytilineos J, Scherer S, Opelz G: Comparison of RFLP-DR
beta and serological HLA-DR typing in 1500 individuals. Transplantation
50:870, 1990 [PMID
1978425]
-
Mytilineos J, Lemoert M, Middleton D, et al: HLA class
I typing of 215 "HLA-A,-B,-DR zero mismatched" kidney transplants. Tissue
Antigens 50:355, 1997 [PMID
9349619]
-
Bodmer JG, Marsh SGE, Albert ED, et al: Nomenclature
for factors of the HLA system, 1996. Genetic Diversity of HLA: Functional
and Medical Implication, Vol 1. Charron D, Ed. EDK, Paris, 1997, p 505
-
Harding CV, Unanue ER: Cellular mechanisms of antigen
processing and the function of class I and II major histocompatibility
complex molecules. Cell Regulation 1:499, 1990
-
Powis SH, Tonks S, Mockridge I, et al: Alleles and
haplotypes of the MHC-encoded ABC transporters TAP1 and TAP2. Immunogenetics
37:373, 1993 [PMID
8428770]
-
Hunt DF, Michel H, Dickinson TA, et al: Peptides presented
to the immune system by the murine class II major histocompatibility complex
molecule I-Ad. Science 256:1817, 1992 [PMID
1319610]
-
Chicz RM, Urban RG, Gorga JC, et al: Specificity and
promiscuity among naturally processed peptides bound to HLA-DR alleles.
J Exp Med 178:27, 1993 [PMID
8315383]
-
Von Boehmer H: Positive selection of lymphocytes. Cell
76:219, 1994
-
Garboczi DN, Ghosh P, Utz U, et al: Structure of the
complex between human T-cell receptor, viral peptide and HLA-A2. Nature
384:134, 1996 [PMID
8906788]
-
Sayegh MH, Watschinger B, Carpenter CB: Mechanisms
of T cell recognition of alloantigen: the role of peptides. Transplantation
57:1295, 1994 [PMID
8184464]
-
Fangmann J, Dalchau R, Fabre JW: Rejection of skin
allografts by indirect allorecognition of donor class I major histocompatibility
complex peptides. J Exp Med 175:1521, 1992
-
Sayegh MH, Khoury SJ, Hancock WW, et al: Induction
of immunity and oral tolerance by polymorphic class II major histocompatibility
complex allopeptides in the rat. Proc Natl Acad Sci USA 89:7762, 1992 [PMID
1502196]
-
Matsushita S, Muto M, Suemura M, et al: HLA linked
nonresponsiveness to Cryptomeria japonica pollen antigen: I. Nonresponsiveness
is mediated by antigen-specific suppressor T cells. J Immunol 138:109,
1987
-
Thorsby E: HLA-associated disease susceptibility: which
genes are involved? The Immunologist 3:51, 1995
-
Todd JA, Acha-Orbea, Bell JI, et al: A molecular basis
for MHC class II-associated autoimmunity. Science 240:1003, 1988 [PMID
3368786]
-
Thorsby E: HLA-associated diseases. A summary of the
12th International Histocompatibility Workshop component. Genetic Diversity
of HLA: Functional and Medical Implication, Vol 2. Charron D, Ed. EDK,
Paris, 1997, p 91
-
Sollid LM, Marjussen G, Ek J, et al: Evidence for a
primary association of celiac disease to a particular HLA-DQ a/b
heterodimer. J Exp Med 169:345, 1989 [PMID
2909659]
-
Abbas AK, Lichtman AH, Pober JS: The major histocompatibility
complex. Cellular and Molecular Immunology, 2nd ed. WB Saunders Co, Philadelphia,
1994, p 96
-
Carpenter CB: The major histocompatibility gene complex.
Harrison's Principles of Internal Medicine, 11th ed. Braunwald E, Isselbacher
KJ, Petersdorf RG, et al, Eds. McGraw-Hill Book Co, New York, 1987, p 341