Inmunología
⭳ Abrir artículo (PDF)1.1 MBEste artículo fue revisado respecto a la Edición 3/2000. Ver esa versión →
Contenido del artículo
III MECANISMOS DE LA RESPUESTA INMUNE
- Interacciones célula-célula
- Procesamiento y presentación del antígeno a las células T
- UNION NORMAL DEL PEPTIDO DENTRO DE LAS MOLECULAS DEL CPH DE CLASE I Y CLASE II
- UNION DEL SUPERANTIGENO CON LAS MOLECULAS DE CLASE II
- PROCESAMIENTO DEL ANTIGENO
- CELULAS PRESENTADORAS DE ANTIGENO PROFESIONALES
- PRESENTACION DEL ANTIGENO A LAS CELULAS T
- MOLECULAS DE ADHESION
- Transducción de señales en las células T al reconocer al antígeno
- Citocinas y receptores de citocinas
- INTERLEUCINA-2
- INTERLEUCINA-15
- INTERLEUCINA-4
- INTERLEUCINA-13
- INTERFERON GAMA
- FACTOR DE TRANSFORMACION DE CRECIMIENTO-ß
- INTERLEUCINA-10
- INTERLEUCINA-12
- INTERLEUCINA-5
- INTERLEUCINA-6
- INTERLEUCINA-7
- INTERLEUCINA-14
- FACTOR INHIBITORIO DE LA MIGRACION
- FACTOR DE NECROSIS TUMORAL- a
- LINFOTOXINA
- INTERLEUCINA-1
- INTERLEUCINA-8 Y OTRAS QUIMOCINAS
- LINFOTACTINA
- FACTOR ESTIMULADOR DE COLONIAS DE GRANULOCITOS Y MACROFAGOS
- Papel de los carbohidratos en la regulación de las respuestas de las células T
- Activación de las células B
- Inmunidad mediada por células
III MECANISMOS DE LA RESPUESTA INMUNE
DR. JOHN DAVID
DR. COX TERHORST, PH.D.
Interacciones célula-célula
Cuando las células del sistema inmune encuentran un antígeno tiene, lugar una respuesta inmune humoral, celular o ambas. Las respuestas inmunes dependen de linfocitos específicos. La inmunidad humoral es mediada por los linfocitos B, los cuales después de la estimulación proliferan y se diferencían en células plasmáticas productoras de anticuerpos. La inmunidad celular es mediada por los linfocitos T, que al ser activados secretan varias sustancias importantes en la respuesta inmune y que inician diversas reacciones. La primera etapa que conduce a la activación de las células efectoras de ambas divisiones del sistema inmune, consiste en la interacción del antígeno sobre una célula portadora de antígenos (CPA) con el linfocito T cooperador. Los linfocitos B y T se comunican entre sí a través de interacciones que implican receptores (v.gr., interacción entre el receptor de células T [RCT] y el antígeno o la interacción entre varias moléculas de adherencia celular y sus ligandos) y mediante la secreción de varios factores solubles denominados citocinas.1
Procesamiento y presentación del antígeno a las células T
El repertorio de las células T se caracteriza por receptores de superficie celular cuya población muy diversa de regiones variables (V) facilita respuestas muy específicas ante los antígenos. Las células T prototipo ab+ tienen un RCT formado por cadenas a y b (RCT-ab). Estas células responden a antígenos no propios solo cuando los RCT-ab se ponen en contacto con derivados de péptidos antigénicos unidos a moléculas del complejo principal de histocompatibilidad (CPH) localizadas en la superficie celular, un proceso denominado restricción del CPH. El reservorio diverso de las células T se desarrolla conforme las células T inmaduras son sometidas a restricción del CPH dentro del timo durante su desarrollo temprano, y en ese momento son seleccionadas en forma positiva o negativa según su avidez diferencial a antígenos. Si se seleccionan en forma positiva, los linfocitos T con restricción a moléculas del CPH de clase I se convierten en células CD8+, y las que tienen restricción a moléculas de clase II del CPH se convierten en linfocitos CD4+.
Una fracción variable de células T periféricas gdt, tienen un RCT formado por cadenas g y d (RCT-gd), para el que no se conoce hasta el momento un elemento de restricción. Al parecer las células T gdt no requieren presentación del antígeno por las moléculas del CPH.
UNION NORMAL DEL PEPTIDO DENTRO DE LAS MOLECULAS DEL CPH DE CLASE I Y CLASE II
Los productos del CPH son glucoproteínas de superficie celular que unen péptidos antigénicos (i.e., antígenos procesados) dentro de una célula (v.gr., un macrófago) y los transportan a la superficie celular para la interacción con células T [ver adelante, Procesamiento del antígeno]. En el CPH existe un gran polimorfismo (i.e., un gran número de alelos por locus). Sin embargo, dentro de la población humana, cada individuo expresa solo un pequeño número de proteínas diferentes del CPH. Para asegurar una respuesta inmunológica adecuada contra un gran número de antígenos no propios, cada proteína del CPH debe ser capaz de unirse a gran cantidad de péptidos diferentes.
La proteínas de clase I del CPH consisten en una cadena pesada a y una cadena ligera b2-microglobulina (b2-M), mientras que las proteínas de clase II están formadas por cadenas a y b de casi el mismo tamaño [ver la figura 1].2 Las dos moléculas comparten una estructura semeante: dos dominios parecidos a las inmunoglobulinas y un sitio de unión del péptido formado por una hoja de ocho cadenas con plegamiento b y dos regiones a-helicoidales . Mientras que las proteínas de clase I del CPH unen solo péptidos pequeños de longitudes definidas, las proteínas de clase II unen péptidos más grandes, sin aparente restricción por su longitud. Un dato interesante es que ciertos péptidos se unen solo a alelos específicos de las moléculas del CPH de clase I o clase II, por lo que las personas que no tienen el alelo específico no desarrollarán una respuesta inmunológica al péptido asociado.
Con base en estudios de cristalografía y fisicoquímicos se ha propuesto un mecanismo común para la unión de los péptidos a las moléculas de clase I del CPH: los péptidos, generalmente formados por residuos de ocho a 11 aminoácidos de longitud, se unen en el extremo aminoterminal y carboxiterminal al sitio de unión en la molécula de clase I del CPH por medio de una red de uniones hidrógeno a los residuos aminoácidos conservados. Los péptidos más largos se acomodan enrollándose en el centro [ver figura 1a].
La unión de los péptidos a las moléculas de clase II sigue distintos principios.3,4 Estos péptidos tienden a ser mayores y de longitud artribraria. Los residuos aminoácidos conservados forman uniones de hidrógeno en todo el sitio de unión, asegurando la cadena principal del péptido en gran parte de su longitud [ver figura 1b].
Las diferencias en la unión de péptidos entre las proteínas del CPH de clase I y II son causadas por una serie de diferencias estructurales pequeñas dentro de un marco del sitio de unión del CPH relativamente fijo. Estos dos modos de unión probablemente se explican por diferencias fundamentales en el procesamiento de los péptidos que tiene lugar en el retículo endoplásmico (RE) para las moléculas de clase I del CPH y en los endosomas y lisosomas para las moléculas de clase II [ver adelante, Procesamiento del antígeno]. Independientemente de estas diferencias, el área de superficie del péptido central disponible para el reconocimiento directo por un RCT (400 a 500 Å2) es aproximadamente igual para las moléculas de clase I y de clase II.
UNION DEL SUPERANTIGENO CON LAS MOLECULAS DE CLASE II
Un método alternativo de presentación de antígeno incluye a las moléculas de clase II del CPH y a una familia de proteínas que se denominan superantígenos.5 Los superantígenos constituyen una clase de moléculas inmunoestimuladoras asociadas con enfermedades que se unen a moléculas de clase II y estimulan linfocitos T. Los superantígenos han sido implicados en las enfermedades virales, como rabia, síndrome de inmunodeficiencia adquirida (SIDA) y tumores mamarios murinos, y en respuestas de células T a toxinas bacterianas (v.gr., Staphylococcus aureus en el síndrome de choque tóxico y en la intoxicación alimentaria). Aún más, los superantígenos pueden causar enfermedades autoinmunes, como artritis reumatoide. Estudios químicos de las proteínas y cristalográficos han dado evidencias de que los superantígenos se unen a las moléculas de clase II por fuera del sitio convencional de fijación de antígeno. Por ejemplo, el superantígeno bacteriano intacto constituido por la enterotoxina B de S. aureus se une al dominio a1 de la molécula de clase II del CPH humano HLA-DR1. Por lo tanto, se crea un nuevo sitio de fijación al RCT adyacente al sitio de unión convencional del CPH.Aunque los péptidos antigénicos no inhiben la estimulación de las células T por los superantígenos unidos al CPH, los superantígenos pueden bloquear las interacciones convencionales péptido-molécula del CPH con el RCT por obstrucción estérica. Los diferentes alelos de clase II del CPH tienen distintas constantes de unión para superantígenos, causando activación de un repertorio diferente de células T.
PROCESAMIENTO DEL ANTIGENO
Moléculas de clase I del CPH
Las moléculas de clase I del CPH presentan péptidos derivados de proteínas nucleares y citosólicas endógenas degradadas, así como proteínas introducidas por patógenos intracelulares. Para llegar al sitio de unión del péptido de la molécula de clase I del CPH en la luz del RE, los péptidos tienen que cruzar la bicapa lípida del RE. Un sistema de bomba de transporte de péptidos dependiente de trifosfato de adenosina (ATP), que consiste en dos proteínas denominadas TAP1 y TAP2, realiza esta función indispensable. Los requerimientos de esta bomba para pasar el péptido al RE pueden medirse con facilidad porque el ensamble de la cadena pesada a y de la cadena ligera b2M de la molécula de clase I depende de la presencia del péptido. Si no se forma el completo ab2M-péptido, las moléculas de clase I no viajan hasta la superficie celular y son degradadas en la luz del RE. Debido a que las proteínas TAP son polimóficas, aumentan la selectividad de las interacciones péptido-molécula del CPH.
Dos sistemas proteolíticos presentes en todas las células nucleadas se utilizan para atraer a los péptidos hacia la luz del RE para su presentación a las moléculas de clase I. El primero, conjugación ubicua seguida de degradación proteolítica en los proteosomas (subunidades que contienen muchas peptidasas) parece ser critico en la generación de la mayoría de los péptidos presentados [ver figura 2a].6 Sin embargo, el origen preciso de los péptidos no es claro, y pueden originarse en cualquier sitio dentro del citosol. También proteínas no ubicuas y de vida media prolongada pueden ser degradadas en el citosol por los proteosomas. En el segundo mecanismo, antígenos como las proteínas de las cubiertas virales se secretan en la luz del RE, en donde sufren degradación proteolítica por un grupo de proteasas residentes. Como en el caso de los proteosomas, estas proteasas del RE están diseñadas para eliminar de la célula moléculas proteicas sintetizadas en exceso y que no están ensambladas o con constitución inadecuada.
Moléculas de clase II del CPH
Cuando una proteína es introducida a la célula a través de endocitosis, puede ser degradada en los endosomas, lisosomas o en un subcompartimiento endosómico aún no bien definido. Es en estos compartimientos en los que tiene lugar el ensamble entre la molécula de clase II del CPH y el antígeno7,8 [ver figura 2b]. Las proteínas pueden ser procesadas a través de proteólisis antes de la interacción con las moléculas de clase II o, en forma alterna, proteínas antigénicas parcialmente desdobladas pueden unirse a las moléculas de clase II del CPH antes de sufrir degradación proteolítica. Esto puede explicar porqué la longitud de los péptidos unidos a las moléculas de clase II del CPH puede ser muy variable. Después de que el péptido se ha unido a la molécula de clase II del CPH, el endosoma que contiene este complejo migra a la superficie celular y se fusiona con la membrana plasmática.
Aunque la degradación semejante al proteosoma no participa en el procesamiento de los péptidos que se unen a las moléculas de clase II del CPH, dos de los elementos reguladores claves ayudan a controlar las interacciones péptido-molécula de clase II: la cadena invariante y el gen HLA-DM. La cadena invariante, o Ii, bloquea el surco del péptido durante el ensambre de las subunidades a y b de las moléculas de clase II del CPH, evitando así la unión de péptidos procesados por el RE. Es degradada o eliminada del complejo en el endosoma, lo que permite al péptido ocupar el sitio de unión. Se ha demostrado que el gen HLA-DM codifica una función accesoria en el ensamble del complejo molécula clase II del CPH-péptido.9-11 El análisis de las formas mutantes de HLA-DM indican que este producto genético (una proteína semejante a la cadena ? de clase II del CPH) es indispensable en la formación del complejo.
Otras moléculas
Un tercer grupo de moléculas presentadoras de antígeno son las proteínas no polimóficas parecidas a las de clase I asociadas a la b2M. Estas incluyen los productos de los genes no polimóficos CD1 (Cd1a, CD1b, CD1c y CD1e) que se localizan en el cromosoma 1, en lugar de en el cromosoma 6 como las del CPH. Se ha demostrado que CD1b presenta lípidos del ácido micólico de Mycobacterium tuberculosis a células T ab+.12 Este ejemplo de reconocimiento de antígeno no proteico microbiano sugiere que las células T ab+ reconocen un rango de antígenos más amplio de lo que antes se pensaba.
CELULAS PRESENTADORAS DE ANTIGENO PROFESIONALES
Aunque las moléculas de clase I del CPH se expresan en la superficie de todas las células eucariotas, las moléculas de clase II tienen una distribución tisular más restringida. De hecho, ciertas células, incluyendo linfocitos B, células dendríticas, macrófagos, células de Langerhans y células endoteliales, se denominan células presentadoras de antígeno profesionales porque presentan péptidos antigénicos junto con moléculas de clase II del CPH con más eficacia que otras CPA. Esta eficacia se atribuye principalmente a su capacidad para procesar antígenos endocitados. Las CPA profesionales también interactúan con más eficacia con las células T porque tienen varios marcadores de superficie celular, como B7, que se fija a contraestructuras de la superficie de los linfocitos T. Por lo tanto, con frecuencia se realiza una distinción operativa entre las CPA profesionales y no profesionales.
PRESENTACION DEL ANTIGENO A LAS CELULAS T
La detección de complejos molécula del CPH-péptido por las células T ab+ es un proceso dinámico que incluye interacciones intercelulares complejas antes de que tenga lugar la activación del linfocito T. El reconocimiento del antígeno por la célula T sucede en dos fases distintas: (1) un paso de adhesión no específica antes del reconocimiento antigénico, seguido por (2) una interacción específica entre el RCT y el complejo antígeno-molécula del CPH. Este proceso incorpora dos propiedades fundamentales del linfocito T: su capacidad para migrar por todo el organismo y adherirse a muchos tipos de células y su gran especificidad por antígenos particulares.
Las CPA pueden procesar en forma simultánea muchos antígenos proteicos y pueden expresar en su superficie un gran número de complejos diferentes antígeno-molécula del CPH. Solo un número pequeño de estos complejos puede ser reconocido por un determinado linfocito T ab+. A través de la adhesión intercelular, las células T buscan a las CPA potenciales. La adhesión se aborta en ausencia de reconocimiento específico del antígeno por el RCT y se intensifica cuando se logra el contacto correcto RCT-molécula del CPH-péptido. Al adherirse, los complejos RCT-CD3 se agregan en la superficie de membrana de la célula T y se unen a complejos molécula CPH-péptido que se han agregado en la superficie de la CPA.
El orden de los eventos en la búsqueda célula T-CPA se estudió por primera vez en el caso de la función efectora de las células T citotóxicas.13 La fase de adhesión célula T citotóxica-célula blanco dependiente de Mg+2 es seguida de una fase dependiente de Ca+2 que causa la entrega de químicos citotóxicos a la célula blanco. La célula T citotóxica se disocia entonces de la célula blanco, y ocurre muerte de la célula blanco en ausencia de la célula T citotóxica, que recicla para atacar a otra célula blanco. Si la célula T citotóxica que se adhiere a la célula no porta la combinación molécula del CPH-péptido correcta, no se liberan químicos citotóxicos y las células se disocian con más rapidez.
La adhesión de células citotóxicas T a células blanco causa activación de las células T citotóxicas y rearreglo extenso de las estructuras en el citoplasma de la célula T citotóxica. El centro organizador de microtúbulos y el complejo de Golgi se reorientan hacia el sitio de contacto de la célula T citotóxica y la célula blanco. También se acumulan componentes vesiculares en la proximidad del sitio de contacto.
Los anticuerpos monoclonales y las clonas de células T antígeno-específicas fueron cruciales para demostrar que el contacto inicial entre una célula T citotóxica y una célula blanco no requiere de la interacción entre el RCT y su antígeno blanco. Los anticuerpos dirigidos contra las moléculas de adhesión antígeno-1 asociado a la función leucocitaria (LFA-1) y CD2 inhiben las adhesiones tanto específicas como no específicas entre la célula citotóxica y la célula blanco. En forma semejante, anticuerpos monoclonales dirigidos contra las moléculas de adhesión LFA-3 y molécula de adhesión intercelular ICAM-1 bloquean la adhesión a nivel de la célula blanco. Por el contrario, ensayos en agar suave muestran que los rearreglos anticlonotípicos RCT y anti-CD3 no inhiben la adhesión pero sí bloquean la fase lítica. Los anticuerpos anti-CD4 y anti-CD8 inhiben tanto la adhesión como la fase lítica.
Las interacciones entre las células T y las CPA tienen el mismo patrón general. Experimentos que emplearon complejos solubles de molécula de clase II del CPH-péptido sugieren que la constante de unión entre el RCT y este complejo es baja.14 Es probable que solo un pequeño número de complejos molécula CPH-péptido se pongan en contacto inicial con el RCT específico en un área de contacto intercelular establecido por la adhesión. En consecuencia, los complejos RCT-CD3 y las moléculas del CPH que portan el péptido correcto migran hacia al región de contacto célula T-CPA. Esto establece una densidad local muy alta de compleos RCT-CD3, que promueve la unión de antígenos y da lugar a activación del linfocito T. La activación causa reclutamiento de las moléculas de transducción intracelular y un aumento temporal en la adhesión. La activación causa relutamiento de moléculas de transducción intracelular y un aumento temporal en la adhesión. La menor regulación de las moléculas de adhesión permite que la célula se libere. En ausencia de reconocimiento del antígeno, los complejos RCT-CD3 son incapaces de acumularse y ocurre liberación inmediata.
Varias observaciones recientes apoyan este modelo. Los complejos RCT-CD3 tienen propensión a formar agregados en la superficie de la célula T, mientras que las moléculas de clase II del CPH forman dímeros en la superficie de la CPA. Se ha demostrado que se encuentran correceptores CD8 en la superficie de linfocitos T humanos y timocitos como dímeros y multímeros. Por lo tanto, es probable que la interacción receptor-ligando en la interfase del linfocito T con la CPA cause agregación del receptor.
MOLECULAS DE ADHESION
Las principales moléculas de adhesión en las células T son CD2 y LFA-1, que está formada por una cadena alfa (o CD11a) y una cadena b (o CD18).15,16 El ligando natural en la superficie celular para CD2 es LFA-3, o CD58, mientras que el ligando de LFA-1 es ICAM-1 o CD54. Estos ligandos se expresan en todas las CPA potenciales [ver tabla 1]. CD4 y CD8 constituyen una categoría diferente en el paso de adhesión porque, al igual que el RCT, sus ligandos naturales son las glucoproteínas de superficie de clase II y clase I respectivamente del CPH. Las interacciones directas entre CD8 y las moléculas de clase I y entre CD4 y las de clase II tienen un papel crucial en la selección positiva durante la maduración intratímica de los linfocitos T [ver figura 3]. El fenotipo de una célula T determina su función efectora: las células T que expresan CD4 reconocen a las moléculas de clase II del CPH, mientras que las células T que expresan CD8 tienen afinidad por moléculas de clase I. Las moléculas de adhesión celular de los linfocitos T (v.gr., CD2, CD4, CD8 y LFA-1) tienen también una función en la transducción de señales y en la activación de la célula T, que causa una mayor avidez en las interacciones RCT-ligando. A su vez, la transducción de la señal del RCT aumenta la afinidad entre las moléculas de adhesión LFA-1 e ICAM-1.
CD28, un homodímero asociado a disulfuros formado por dos cadenas glucoproteicas de 44kd, es miembro de la superfamilia de genes de inmunoglobulina, específicamente es miembro de la subfamilia de moléculas que tienen un solo dominio V. CD28 se expresa en todas las células T CD4+ y en la mayoría de las CD8+. Una estructura complementaria, denominada B7-1 (o también CD80) se expresa en menor densidad en células B en reposo y puede facilitar la activación de los linfocitos B. B7-1 puede ser inducido en la superficie de las CPA profesionales, como los macrófagos y las células dendríticas.
Otro antígeno de las células T, CTLA-1, se une a B7-1 con mayor actividad y es homólogo a CD28. Se ha descrito una segunda estructura complementaria para CD28 y CTLA-4 en ratones (B7-2 o CD86) y en humanos (B70 o CD86). El perfil de expresión de B7-2 es semejante al de B7-1, con la importante excepción de que B7-2 se expresa en células dendríticas y monocitos en reposo. Se ha demostrado que ratones con deficiencia de CD28 tienen menores respuestas inmunológicas dependientes de células T, lo que demuestra la importancia de este ligando en la activación de las células T.
Transducción de señales en las células T al reconocer al antígeno
En respuesta al reconocimiento del antígeno, las células T en reposo sufren una serie de eventos complejos conocidos como activación.16-18 La señal del RCT no es suficiente per se para causar que las células T en reposo secreten cantidades máximas de interleucina-1 (IL-2), una citocina necesaria para la proliferación de las células T. Por ejemplo, las células T purificadas no producen IL-2 en respuesta a los anticuerpos anti-RCT en ausencia de células accesorias viables. Además, los linfocitos B activados y en reposo tienen capacidad muy diferente para estimular a las células T incluso si el número de complejos molécula CPH-péptido es idéntico. Estas observaciones indican que las CPA hacen más que presentar al antígeno, también proporcionan señales coestimuladoras. Las moléculas de adhesión de las células T CD4, CD8, y CD28 y CTLA-4 también proporcionan señales coestimuladoras importantes.
SEÑALES DEL RECEPTOR DE LA CELULA T
Se han identificado varios eventos bioquímicos que ocurren inmediatamente después del reconocimiento del antígeno por la célula T. El primero parece ser el reclutamiento de cinasas de tirosina-proteína por los elementos CD3-e y CD3-z del complejo RCT-CD3 y quizá por los elementos CD3-g y CD3-d. Esto causa fosforilación tirosina de las colas citoplásmicas de CD3-s y CD3-e y, en menor grado, de CD3-g y CD3-d. Dos tirosina cinasas principales, p56lck y p59fyn (ambas relacionadas con la familia del proto-oncogén src), participan en la tirosina fosforilación de CD3. El nivel de fosforilación de CD3-s depende de la naturaleza del complejo molécula CPH-péptido reconocido por el RCT-ab.
Cuando se une al complejo RCT-CD3, la p59fyn fosforila y activa a la enzima celular fosfolipasa C-g (PLC-g), que induce la liberación de Ca2+ de las reservas intracelulares al generar inositol 1,4,5-trifosfato (IP3). Esta liberación de Ca2+ es la característica de la activación celular en muchos sistemas, incluyendo las células T y B. Durante el reconocimiento del antígeno, la fosforilación de las colas citoplásmicas CD3-e y CD3-s causan reclutamiento de dos tirosina cinasas diferentes, p72syk y ZAP-70, al complejo RCT-CD3. Las proteínas blanco precisas para p72syk y ZAP-70 se desconocen, pero pueden incluir moléculas inhibitorias de transducción de señales, como p95vav y p72-74raf [ver figura 4].
El ciclo de fosforilación-defosforilación de tirosina es necesario para la activación de la célula T. Las cinasas de tirosina son vitales en estas reacciones. Se ha demostrado que los inhibidores de estas enzimas bloquean las fases tempranas de activación de las células T, mientras que la depleción de la proteína cinasa C (PCC) no lo hace. También es importante la presencia de CD45, una fosfatasa tirosina unida a la membrana. La ausencia de CD45 en las células T inhibe la vía de activación en varios grados, la reconstitución de CD45 por transfección con ADN complementario (ADNc) de CD45 restablece su actividad total. Se sabe que la cinasa carboxi terminal src (csk) inhibe la producción de IL-2 inducida por el RCT en modelos experimentales: csk fosforila los sitios reguladores inhibitorios en las cinasas relacionadas a src p59fyn y p56lck, inactivando estas enzimas. La importancia de la fosforilación de tirosina en la activación de la célula T ha sido demostrada por estudios de ratones mutantes en los que se han alterado los genes fyn, lck o CD45. En otro estudio, los pacientes que no tenían la cinasa de tirosina ZAP-70 fueron incapaces de reclutar células T CD4+. Sorprendentemente, la ausencia de ZAP-70 causó también ausencia de células T CD8+.19,20
La activación de la PLC-? causa la liberación de diacilglicerol, que a su vez activa a la PCC. La PCC inicia la cascada de otras cinasas de serina-treonina, incluyendo p72-74raf, mek y MAP. La molécula central en la transducción de señales, ras, parece participar en eventos más tardíos, después de la activación de PCC. Tanto la vía ras como las vías de cinasa de serina-treonina están acopladas, porque activan genes tempranos como jun y fos, que están implicados en la activación del gen de la IL-2 [ver figura 4]. El Ca2+ generado por el IP3 activa otra cascada, la vía de la calcineurina. La calcineurina, una fosfatasa de serina-treonina, es una enzima dependiente de calcio y calmodulina que participa en la inducción del factor de transcripciòn NF-AT, que se requiere para la transcripción de IL-2. La vía dela calcineurina es bloqueada por la ciclosporina, que se fija a la ciclofilina, inhibiendo la actividad de la calcineurina.
Otra enzima importante que participa en el metabolismo lípido es la fosfatidilinositol-3 cinasa (PI3-cinasa), una enzima clave en las vías de señales mitogénicas de las cinasas de tirosina tanto del receptor como no de receptor. La PI3-cinasa tiene dos propiedades características: fosforilación de la molécula de inositol por el fosfatidilinositol para producir PI3P y asociaciones específicas con muchas cinasas de proteína activadas. La activación del complejo RCT-CD3 aumenta la actividad de la PI3-cinasa, que se asocia con una cadena fosforilada CD3-s. Aún no se ha determinado si la PI3-cinasa se une en forma directa al CD3-s activado o a través de otras cinasas, como las cinasas de la familia src (v.gr., p56lck y p59fyn) y las cinasas p72syk y ZAP-70.
SEÑALES DE MOLECULAS COESTIMULADORAS
La activación aberrante del RCT puede iniciar respuestas inmunológicas muy dañinas, como las observadas en las enfermedades autoinmunes y otras patologías. Las moléculas coestimuladoras tienen un papel muy importante en la coordinación y regulación cinética de la activación de las células T. En algunos casos las moléculas coestimuladoras de los linfocitos T y sus ligandos en las CPA son inducidas por citocinas y transducción de señales después de la interacción intercelular inicial. Estas moléculas pertenecen a diferentes familias de receptores, incluyendo la superfamilia de las inmunoglobulinas, las superfamilias del factor de necrosis tumoral (FNT) y de los receptores de FNT, las integrinas y las selectivas. Las moléculas coestimuladoras de los linfocitos T y otras moléculas cuya unión puede, en algunos casos, inhibir la activación celular son CD2, CD4, CD5, CD7, CD8, CD27, CD28 y CTLA-4, gp39 (CD40L), CD45, LFA-1 (complejo CD11-CD18), ICAM-1 (CD54) y Fas (CD95) [ver tabla 1].
CD28 y CTLA-1 proporcionan la señal secundaria más importante, iniciando una vía de transducción de señales que es diferente e independiente de la vía mediada por el complejo RCT-CD3. CD28 utiliza una cinasa de tirosina recién descubierta, p72.tsk 21 A diferencia de la señal del complejo RCT-CD3, la señal enviada por CD28 no es bloqueada por ciclosporina. Por lo tanto, la vía de transducción de señales de CD28 puede evitar a la fosfatasa dependiente de Ca2+ calcineurina, lo que apoya el concepto de que esta vía es independiente de Ca2+. Además de tener un papel en la activación de la célula T, la activación de CTLA-4 por B7 provoca la inhibición de la fase tardía de la activación de la célula T. CD28 se expresa en forma constitutiva en la superficie de los linfocitos T, pero CTLA-4, un homólogo de CD28, se expresa solo en los linfocitos T activados. Ni B7-1 ni B7-2 se expresa en forma constitutiva en la superficie de los linfocitos B. Un aspecto que aumenta aún más la complejidad es el hecho de que B7-1 y B7-2 son, per se, capaces de realizar transducción de señales en las células B.
La molécula coestimuladora CD27 se induce al activarse la célula T y actúa en forma sinérgica con la activación asociada al RCT para inducir la proliferación de linfocitos T. CD27 (en los linfocitos T) es un miembro de la familia del receptor del FNT, mientras que CD70, la estructura correspondiente en linfocitos B activados, es codificada por el gen de la superfamilia del FNT.
Otro miembro de la familia del receptor de FNT, Fas (CD95) ha sido implicado en señales tanto positivas como negativas a través de la interacción con el ligando de Fas. La unión del antígeno Fas en las células T típicamente causa muerte celular programada (i.e., apoptosis). Sin embargo, se ha demostrado que la unión con un anticuerpo monoclonal anti-Fas permite que Fas funcione como una molécula coestimuladora para la activación del RCT-CD3. Por lo tanto, una sola molécula puede originar diferentes señales en diferentes estadios del desarrollo del linfocito T.
PAPEL DE CD4 Y DE CD8
Las moléculas CD4 y CD8 son correceptores del RCT que interactúan con las moléculas del CPH en la superficie de las CPA. CD4 es un monómero de 55 kd que consiste en cuatro dominios semejantes a los de las inmunoglobulinas, mientras que CD8 está formado por dos cadenas, a y b, que son codificadas por genes diferentes. CD8-b se expresa solo en unión con CD8-a, pero CD8-a puede presentarse como homodímero. Se ha propuesto que el sitio de unión de CD8 es el dominio alfa3 de la molécula de clase I del CPH. Es probable que el contacto entre CD4 y la molécula de clase II del CPH incluya los primeros tres dominios semejantes a inmunoglobulinas de CD4.
El hecho de que CD4 y CD8 se asocien con la cinasa de tirosina citoplásma p56lck sugiere que participan en la transducción de señales. Se requieren dos residuos de cisteínas en regiones claves de CD4 y CD8-a para la asociación con p56lck. Los anticuerpos anti-CD4 pueden inhibir las respuestas de linfocitos T en ausencia de interacción del RCT con antígenos del CPH, lo que indica que CD4 puede trasmitir también una señal denominada negativa.
La interacción de las moléculas CD4 y CD8 con las moléculas del CPH es indispensable para generar a los linfocitos T CD4+ y CD8+. El cambio de timocitos CD4+, CD8+ (dobles positivos) a CD4+ o CD8+ (positivos únicos) requiere de selección positiva en el timo, que depende de la interacción entre el complejo RCT-CD3 con los antígenos del CPH. El ratón clase Inulo del CPH (obtenido por interrupción dirigida del gen que codifica a la b2M o del gen que codifica a TAP1) tiene un número mínimo de timocitos CD8+ comparado con el ratón normal. En forma semejante, el ratón clase IInulo del CPH no tiene timocitos CD4+. El ratón CD8nulo no posee células T citotóxicas, mientras que el ratón CD4nulo no tiene células T cooperadoras. La falta de células T CD4+ y CD8+ en el ratón CD4nulo y CD8nulo, respectivamente, no fue compensada por el subgrupo celular recíproco. Además, el número de células CD4-, CD8- aumentó, demostrando que se requiere de las moléculas CD4 y CD8 para generar células T CD4+ y CD8+ funcionales.
Los mecanismos de transducción de señales que participan en la selección positiva no se han definido aún. Debido a que también se encontró que la cinasa de tirosina p56lck se asocia con CD4 y CD8 en el timo, existe la posibilidad de que esté involucrada en la selección positiva. Sin embargo, su asociación con CD4 y CD8 no es indispensable, ya que pueden desarrollarse células T CD4+ en ratones con CD4 que no tiene cola citoplásmica, lo que le hace incapaz de asociarse con p56lck. En forma semejante, Loh y colaboradores demostraron en 1993 que la asociación de p56lck con CD8 no era necesaria para el desarrollo de células T CD8+.22
Citocinas y receptores de citocinas
Las citocinas, un grupo diverso de proteínas producidas por varios tipos celulares, son críticas en la regulación de las respuestas inmunológicas.23 También son importantes en la diferenciación de los sistemas celulares. En general, las citocinas no se almacenan en las células, sino que se sintetizan por el estímulo apropiado. El ARN mensajero (ARNm) resultante tiene vida media corta. En algunos casos la citocina activa es liberada a partir de un precursor inactivo por proteólisis. Después de su secreción, las citocinas tienen acción local, pueden actuar sobre muy diferentes células, y distintas citocinas pueden tener actividades semejantes. Los receptores de citocinas comparten también cadenas proteicas comunes.24 Por ejemplo, los receptores de citocinas IL-2R, IL-4R, IL-7R, IL-9R, IL-13R e IL-15R comparten cadenas g comunes.25,26 La cadena polipeptídica gp130 es compartida por los receptores de la IL-6, el factor inhibidor de leucemia (FIL) y el factor neurotrópico ciliar (CNTF). Los receptores para IL-3, IL-5 y el factor estimulador de colonias de granulocitos y macrófagos (FEC-GM) comparten una cadena b en común [ver tabla 2]. La mayoría de las citocinas forman una red que regula la activación de las células y la producción de otras citocinas.
Las citocinas tienen un papel importante en el desarrollo y regulación de las células T, De hecho, los subgrupos TH1 y TH2 se definen por las citocinas que producen.
Los linfocitos TH1 se desarrollan bajo la influencia de la IL-2 y el interferón gama. Producen interferón gama, IL-2 y FNT-alfa y participan principalmente en las reacciones de inmunidad mediada por células e hipersensibilidad. El interferón gama es el factor activador más importante para los macrófagos.
Las células TH2 se desarrollan por influencia de IL-4 y participan sobre todo en reacciones alérgicas, producción de anticuerpos y cambio de clase de anticuerpo (en especial de IgE a IgA). Estas células producen IL-4, IL-5, IL-6 e IL-10. La IL-10 inhibe la activación de los macrófagos, lo mismo que la IL-4. Cada uno de estos dos subgrupos de linfocitos T cooperadores inhibe el desarrollo y función del otro. El interferón gama producido por las células TH1 inhibe el desarrollo y función de las células TH2. Por el contrario, la IL-4 y la IL-10 producidas por las células TH2 inhibe el desarrollo y función de los TH1 [ver figura 5]. La IL-12 aumenta la función de las células TH1 porque es un potente factor de crecimiento para las células asesinas naturales (NK), que también producen interferón gama.
El hecho de que las células TH2 inhiban las respuestas inmunológicas mediadas por TH1 y que las TH1 inhiban las respuestas mediadas por anticuerpos de las TH2 puede atribuirse a que ambos subgrupos inducen también inflamación, que debe controlarse [ver adelante, Reacciones de hipersensibilidad tardía].
INTERLEUCINA-2
La IL-2 es producida por las células T después de la estimulación por un antígeno o mitógeno, y es el principal factor de crecimiento para las células T CD4+ y CD8+. También influye en el desarrollo de los linfocitos B. La IL-2 estimula el crecimiento y la función citolítica de células NK específicas conocidas como células activadas por linfocinas (LAK). Dos retrovirus, el virus linfotrópico de células T humanas tipo I (HTLV-I) y el HTLV-II, producen una proteína transactivadora que estimula la expresión de los genes que codifican tanto para la IL-2 como para su receptor. Los anticuerpos monoclonales contra el receptor de la IL-2 tienen un papel potencialmente terapéutico en la anemia aplástica y la leucemia causada por HTLV-I.
La IL-2 es una glucoproteína de 15 a 17 kd (heterogénea por glucosilación diferente). Es miembro de la familia de citocinas hematopoyéticas y, como todos los miembros, está compuesta de cuatro segmentos de alfa-hélice organizados en dos haces paralelos. La IL-2, la IL-4 y el FEC-GM tienen una estructura muy semejante, y se unen a receptores similares.27 El gen que codifica para la IL-2 se localiza en el cromosoma humano 4. La actividad de la IL-2 no es especie específica.
Se ha demostrado la importancia de la IL-2 por estudios de ratones mutantes con gen de la IL-2 alterado (ratón IL-2nulo): la mitad de estos ratones mueren por anemia severa causada por autoanticuerpos dirigidos contra eritrocitos.28 Los que no mueren por anemia lo hacen por colitis ulcerativa a menos que se mantengan en un ambiente libre de gérmenes. La IL-2 puede ser importante en la eliminación de células autorreactivas.29 Los ratones en los que se alteró el gen que gobierna la cadena ? del receptor de IL-2 (ratones IL-2Rbnulo) murieron por anemia severa a menor edad que los ratones IL-2nulo. La diferencia entre los dos ratones mutantes radica en el hecho de que la IL-15 aún puede funcionar en el IL-2nulo, pero no en el IL-2Rbnulo, porque la IL-2Rb también es parte del receptor de la IL-15.
INTERLEUCINA-15
La IL-15, recién aislada del sobrenadante de cultivos de líneas celulares de epitelio de riñón de simios, apoya la proliferación de una línea celular dependiente de IL-2.30 Estimula a las células que usan las cadenas b y g del receptor de la IL-2, pero no a las que usan la cadena a. Los anticuerpos contra IL-2R bloquearon la actividad de la IL-15. Esta citocina es una proteína de 14 a 15 kd que no tiene homología en su secuencia de aminoácidos con ninguna otra citocina. Un factor semejante de 14kd, denominado como IL-T, también tiene propiedades semejantes a la IL-2 que estimulan la proliferación de las células T e inducen la formación de células LAK. Se ha encontrado que la IL-T no es bloqueada por anticuerpos contra IL-2 o IL-2Ra, pero sí por anticuerpos contra IL-2Rb e IL-2Rg. Las consecuencias de que la cadena polipéptida gama es compartida en los receptores para IL-2, IL-4, IL-7, IL-9, IL-13 e IL-15 se han demostrado ampliamente en pacientes con inmunodeficiencia combinada severa (IDCS) ligada al cromosoma X, que es causada por falta de la cadena polipeptídica gama.
INTERLEUCINA-4
La IL-4, producida principalmente por linfocitos T, es crucial en el desarrollo del subtipo de células TH2 a partir de las células precursoras TH0.23 Las células TH2, a su vez, producen IL-4 adicional, así como IL-5, IL-6, IL-10 e IL-13. Estas células participan en la ayuda a linfocitos B, y la IL-4 es la principal citocina que causa el cambio en la producción de anticuerpos a IgE, y en menor grado a IgG1, en las células B. La IL-4 es un factor de crecimiento para células cebadas y estimula a la molécula de adhesión de células vasculares-1 (VCAM-1) sobre las células endoteliales, aumentando su capacidad para fijar eosinófilos, linfocitos y monocitos. Las células cebadas activadas y los basófilos producen IL-4 adicional. Las células TH2 son importantes para mediar las reacciones alérgicas, así como la inmunidad a nemátodos, en la que los anticuerpos tienen un papel muy importante. Por otro lado, la IL-4 inhibe el desarrollo del subgrupo TH1 y los efectos activadores sobre macrófagos del interferón gama, incluyendo la actividad microbicida y la producción de IL-1 y de óxido nítrico. Por lo tanto, la IL-4 inhibe la inmunidad protectora contra organismos intracelulares y suprime los procesos inflamatorios inducidos por las células TH1.
La IL-4 es una glucoproteína de 20 kd miembro de la familia de citocinas hematopoyéticas. Se codifica en el cromosoma humano 5, cerca del gen de la IL-5. La actividad de la IL-4 es muy específica de especie. El ratón IL-4 nulo tiene alteración en el cambio de isotipo de inmunoglobulinas y células TH2 parcialmente disfuncionantes.31,32 La IL-13 tiene la capacidad de realizar algunas de las funciones de la IL-4.33
INTERLEUCINA-13
La IL-13, producida por las células T CD4+ y CD8+ activadas, comparte muchas de las funciones de la IL-4.33 Actúa sobre los macrófagos para suprimir varias citocinas proinflamatorias, quimocinas (i.e, citocinas quimiotácticas) y factores de crecimiento hematopoyético (incluyendo IL-1, IL-8, FNT-a, proteína inflamatoria de macrófagos-1a [PIM-1a] y FEC-GM), inhibe la citotoxicidad mediada por células dependiente de anticuerpos (CMCDA) al disminuir el número de receptores Fc, e inhibe la producción de óxido nítrico en macrófagos murinos, y con ello la capacidad de los macrófagos para destruir parásitos intracelulares, como Leishmania. Sin embargo, la IL-13 aumenta la presentación de moléculas clase II del CPH y antígenos. A diferencia de la IL-4, la IL-13 actúa sobre los linfocitos B humanos para cambiar la producción de anticuerpos a IgE e IgG4 (pero no lo hace en linfocitos B murinos). Aunque diferentes, los receptores IL-4R e IL-13R comparten algunas características. El cambio de un aminoácido en la IL-4 provoca una molécula que se ha denominado hIL-4.Y124D, que no tiene la actividad de la IL-4 pero aún se une al receptor e inhibe la acción tanto de IL-4 como de IL-13. La capacidad de esta IL-4 mutada para inhibir la síntesis de IgE por la IL-4 y la IL-13 puede ser útil en algunas estrategias de tratamiento.
La IL-13 es una proteína de 10 kd que comparte alrededor del 30 porciento de aminoácidos homólogos con la IL-4. Se transcribe antes que la IL-4 en los linfocitos T activados y durante un periodo más prolongado.
INTERFERON GAMA
El interferón gama, también llamado tipo II o interferón inmune, es producido por las células T y NK. Es crucial en el desarrollo de los linfocitos TH0 a TH1 e inhibe el desarrollo de células TH2. Es el principal factor activador de macrófagos (FAM) y también es una potente citocina en la inmunidad mediada por células. Otros FAM incluyen al FEC-GM y al factor inhibidor de la migración (FIM). La IL-1 y el FNT-a tienen también leve actividad de FAM. La IL-12 estimula a las células NK para producir mayores cantidades de interferón gama, aumentando las reacciones dependientes de esta citocina. El interferón gama aumenta la expresión de moléculas de clase I y clase II del CPH, aumentando la presentación de antígenos y otras reacciones cognoscitivas. Per se y por los mayores efectos del FNT-a, el interferón gama causa la expresión de moléculas de adhesión en la superficie de las células del endotelio vascular., provocando adhesión y extravasación de linfocitos T. El interferón gama se ha empleado con éxito para tratar enfermedades granulomatosas crónicas y leishmaniasis visceral resistente al tratamiento.34
El interferón gama está constituido por dos subunidades de 21 a 24 kd, es heterogéneo por diferencias en la glucosilación. El ratón interferón gamanulo y el ratón con receptor de interferón gamanulo tienen destrucción defectuosa de microrganismos por macrófagos, menor expresión de moléculas de clase II del CPH en los macrófagos y menor producción de óxido nítrico, función deficiente de las células NK y menor concentración sérica de IgG2 e IgG3.
FACTOR DE TRANSFORMACION DE CRECIMIENTO-b
El factor de transformación de crecimiento-b (FTC-b) es producido por diferentes células, incluyendo plaquetas, linfocitos, macrófagos activados y células de la placenta. Está unido en forma no covalente a una proteína asociada de latencia (PAL) de 75 kd, de la cual se libera por acción de una proteasa para adoptar su forma activa. El FTC-b permite a las células normales crecer en agar suave, una característica que suele observarse en células malignas. El FTC-b es una citocina muy pleiotrópica, estimula la producción de proteínas de la matriz extracelular, como la colágena. También cambia a producción de IgA en los linfocitos B (aumentando así las reacciones inmunológicas mucosas) e inhibe la producción de IgM, IgG1 e IgG2. Además, el FTC-b inhibe la activación de macrófagos, la proliferación de linfocitos inducida por mitógenos y la maduración de células T citotóxicas, controlando los efectos de muchas citocinas. De hecho, las reacciones inflamatorias en los ratones FTC-bnulo se controlan muy mal.
El FTC-b está constituido por tres péptidos homodiméricos de 25 kd (FTC-b1, FTC-b2 y FTC-b3) y cada isoforma es codificada por genes diferentes mapeados en distintos cromosomas.
INTERLEUCINA-10
La IL-10 es producida por linfocitos T CD4+ y CD8+, linfocitos B, macrófagos, células cebadas y queratinocitos.36 Aunque casi siempre se asocia con actividad de células TH2, también puede producirse por células TH1.37,38 La IL-10 suprime las respuestas de linfocinas al disminuir la producción de citocinas por los macrófagos, incluyendo IL-1, FNT-a, IL-6, IL-8, FEC-GM, y factor estimulador de colonias de granulocitos (FEC-G), e inhibir la producción de óxido nítrico. A diferencia de su potente actividad inmunosupresora en los macrófagos, la IL-10 estimula a las células B para proliferar, diferenciarse y producir anticuerpos, y también parece ser un factor de cambio para la producción de IgG4.
En humanos, la IL-10 es un polipéptido de 18 kd codificado por un gen en el cromosoma 1. Muestra importante homología en la secuencia de aminoácidos con una proteína del virus Epstein-Barr que tiene cierta actividad de IL-10 in vitro. No se sabe si la IL-10 viral y de mamífero comparten un mismo gen ancestral. La disfunción inmunológica en el ratón IL-10nulo causa enteritis y colitis. La enfermedad puede transferirse en forma adoptiva a los ratones con IDSC por medio de células T purificadas.39
INTERLEUCINA-12
La IL-12 es un potente estimulador de las células NK, causa la maduración de células T citotóxicas y es un factor de crecimiento para las células T activadas. Por su capacidad para inducir el interferón gama, la IL-12 aumenta la formación de células TH1 e inhibe la formación de células TH2 por la IL-4.40,41
La IL-12 puede proteger a los ratones BALB/c susceptibles contra la infección por L. major cuando reciben antígeno de Leishmania antes o después de la infección. Es posible que pueda emplearse esta citocina como adyuvante en el desarrollo de vacunas.42,42
La estructura de la IL-12 es única entre las citocinas, Esta formada por dos subunidades diferentes producidas por distintos tipos celulares. Una subunidad, IL-12a es una proteína de 35 kd producida por células T y B, células NK y macrofagos, Tiene la estructura de cuatro hélices a semejante a otras citocinas hematopoyéticas. La otra subunidad, IL-12b, es una proteína de 40 kd producida por células B y macrófagos activados. Es homóloga al receptor para IL-6, que contiene la secuencia Trp-Ser-X-Trp-Ser. Por lo tanto, parece ser que el heterodímero de IL-12 está constituido por una proteína semejante a una citocina ligada a una proteína semejante a un receptor de citocina.
INTERLEUCINA-5
La IL-5, producida por células TH2 y células cebadas, estimula el crecimiento y diferenciación de los eosinófilos. Los eosinófilos son activados por la IL-5 y por un factor que aumenta la citotoxicidad de los eosinóficos derivado de monoticos. Los eosinófilos activados destruyen parásitos helmintos y producen daño tisular en estados de alergia. La IL-5, antes denominada factor de crecimiento de células B II y factor de remplazo de células T, estimula el crecimiento y diferenciación de los linfocitos B y, en sinergia con otras citocinas, induce la síntesis de IgM e IgA en células B maduras. La IL-5 es un homodímero de 40 kd relacionado con la familia de las citocinas hematopoyéticas.
INTERLEUCINA-6
La IL-6 es producida por diferentes tipos celulares, incluyendo macrófagos, fibroblastos, células vasculares endoteliales y algunas células T activadas, y tiene un efecto muy amplio sobre diversos tipos celulares.44-46 La IL-6, antes denominada factor estimulador de células B-2 y factor de diferenciación de las células B, induce diferenciación terminal de linfocitos B secretores de inmunoglobulinas, Actúa como un factor de crecimiento para las células plasmáticas malignas. De hecho, se ha observado proliferación policlonal extensa de células plasmáticas en ratones transgénicos que expresan un exceso de Il-6. La producción de IL-6 es estimulada por IL-1 y el FNT-a de los macrófagos, pero la IL-6 inhibe la producción por los macrófagos de IL-1 y FNT-a. La IL-6 actúa sobre las células hepáticas para producir proteínas de fase aguda, como fibrinógeno, alfa2-macroglobulina y proteína de amiloide sérico A.
La IL-6, un péptido que varía en tamaño de 21 a 32 kd por la glucosilación, pertenece a la familia de citocinas hematopoyéticas. En el ratón IL-6nulo existe disrregulación de los progenitores mieloides y eritroides. Como es de esperarse, este ratón no produce proteínas de fase aguda y es más resistente a la infección.
INTERLEUCINA-7
La IL-7, un monómero de glucoproteína de 25 kd, es producida por las células del estroma de la médula ósea y del timo y afecta a las células B tempranas. Induce proliferación de los progenitores B (linfocitos B en los que los genes de las cadenas ligeras y pesadas no están aún rearreglados en la configuración de línea germinal) y de células pre-B (que tienen ya rearreglo del gen de la cadena pesada y contienen cadenas pesadas en el citoplasma) pero no de las células B maduras que producen inmunoglobulina asociada a la membrana (i.e., receptores de células B). En este sentido, la IL-7 debe considerarse como relacionada a otras citocinas hematopoyéticas, como IL-3. La IL-7 estimula también el crecimiento de las células T CD3-, CD4- y CD8- que pueden después responder a IL-2. El timo contiene niveles altos de ARNm para IL-7. La IL-7 puede estimular a las células T maduras y se ha demostrado que aumenta las células T citotóxicas. Se observa un aumento importante en las células pre-B de la médula ósea y linfoices en los ratones transgénicos que expresan IL-7 en exceso.
La IL-7 es un monómero de glucoproteína de 25 kd. Los ratones mutantes deficientes en el receptor de IL-7 tienen una reducción muy importante en la celularidad linfoide tímica y periférica.47
INTERLEUCINA-14
La IL-14, un factor de crecimiento de células B humanas producidas por células malignas B y T, también es conocido como factor de crecimiento de células B de alto peso molecular.48 Induce proliferacion de células B activadas, pero no de células B en reposo e inhibe la secreción de inmunoglobulinas. Los estudios preliminares sugieren que expande en forma selectiva la subpoblaciones de células B de memoria normales.48 Cuando se examinan los leucocitos, solo el linaje de células B tiene receptores para IL-14. La IL-14 es una proteína de 53 kd.
FACTOR INHIBITORIO DE LA MIGRACION
El FIM fue la primera citocina de células T que se descubrió, y se produce después de la estimulación de células T sensibilizadas con el antígeno específico. Esta citocina fue el primer parámetro in vitro de las reacciones de hipersensibilidad tardía [ver figura 6]. Los linfocitos pueden estimularse también en forma inespecífica por mitógenos y complejos inmunes para producir FIM. Se ha clonado el gen para el FIM humano y murino, el monómero es una proteína de 12 kd.49 Además, se ha demostrado que el gen para el FIM es un gen temprano que se expresa en muchos tejidos diferentes.50 Se encuentran grandes cantidades del gen en los macrófagos y las células hipofisiarias. De hecho, el lipopolisacárido (LPS) estimula la liberación de FIM por la hipófisis. Cuando se administra a ratones, el FIM aumenta mucho la letalidad del LPS. Por el contrario, los anticuerpos anti-FIM revierten por completo esta mortalidad.51 Esta citocina participa también en el desarrollo del cristalino ocular.52 Se ha demostrado que el FIM recombinante activa a macrófagos para destruir a Leishmania y los estimula para producir FNT-alfa. Aún no se determina la forma como el FIM interactúa con otras citocinas. El FIM parece ser idéntico al factor inhibitorio de glucosilación, que inhibe la producción de IgE.53
FACTOR DE NECROSIS TUMORAL- a
El FNT-a es una de las sustancias más abundantes producidas por los macrófagos, en especial después de la estimulación por endotoxina. Los macrófagos producen también FNT-a después de la estimulación con interferón gama o FIM. El FNT-a es producido por células T activadas, células NK y células cebadas. Su nombre deriva del hecho de que fue un factor que se encontró en el suero de animales que recibían endotoxina que podía inducir necrosis hemorrágica en los tumores. El FNT-a tiene un papel crucial en la respuesta del huésped a bacterias gram negativas, la fuente de la endotoxina y del LPS. El FNT-a media los efectos del LPS, que en dosis bajas induce una respuesta protectora que causa aumento en la función de los macrófagos, poliactivación de linfocitos B y fiebre. la inmunidad protectora de ciertos organismos intracelulares, como Leishmania, aumenta por efecto del FNT-a. Los efectos antivirales de este factor son causados por la producción de interferón gama. Sin embargo, en concentraciones altas el LPS causa muerte por estado de choque (que puede prevenirse por anticuerpos anti-FNT-a) y coagulación intravascular diseminada. Parte de este fenómeno se conoce como la reacción de Shwartzman. El LPS causa la liberación de FIM, que a su vez aumenta la producción de FNT-a por los macrófagos.
En concentraciones bajas, el FNT-a aumenta la respuesta inflamatoria protectora. Esto induce la expresión de moléculas de adhesión ICAM-1, VCAM-1 y selectina E (ELAM-1) en las células endoteliales vasculares, que promueven la adhesión de neutrófilos, macrófagos y eosinófilos. Estimula también la producción de citocinas por los macrófagos, incluyendo el FNT-a en sí. la IL-1, la IL-6 y diversas quimocinas. El FNT-a aumenta la expresión de moléculas de clase I del CPH y potencia la lisis celular inducida por células T citotóxicas. En concentraciones elevadas el FNT-a puede actuar como una hormona. Funciona como un pirógeno semejante a IL-1 (ver adelante), activa el sistema de coagulación y la producción de proteínas de fase aguda hepáticas y puede causar inmunodeficiencia por la supresión de la médula ósea. El FNT-a inhibe la contractilidad del miocardio al estimular la producción de óxido nítrico y, cuando está presente por tiempo prolongado, causa caquexia. Muchos de los síntomas del paludismo, en especial a nivel cerebral, y algunos de los síntomas de la infección por el virus de inmunodeficiencia humana (VIH) pueden ser mediados por FNT-a.
El FNT-a circula en el suero como homotrímero, formado por subunidades de 17 kd que han surgido de proteolisis de un monómero transmembrana de 25 kd. Se ha evaluado tanto a los anticuerpos anti-FNT-a como a los receptores solubles del FNT-a por su posible papel en el tratamiento de diversos padecimientos inflamatorios.
LINFOTOXINA
Producida en forma exclusiva por las células T activadas, la linfotoxina comparte muchas propiedades biológicas con el FNT-a y también se le conoce como FNT-b. Al igual que el FNT-a, la linfotoxina lisa células tumorales pero no células normales, activa neutrófilos y aumenta la adhesión vascular y la extravasación de los leucocitos. Los efectos de la linfotoxina dependen de la unión con otra proteína, la linfotoxina b, para formar un compleo. Utiliza el mismo receptor celular que el FNT-a.
La linfotoxina es una glucoproteína de 21 a 24 kd. Al contrario del FNT-a, es una proteína secretora que no tiene un componente transmembrana. También se conoce la estructura cristalizada de la linfotoxina unida al dominio extracelular del receptor de FNT de 55 kd.54 El complejo tiene tiene tres moléculas receptoras unidades de modo simétrico a un trímero de linfotoxina. El dominio extracelular del receptor, una estructura muy larga y con ensambles término-terminal de cuatro dominios semejantes entre sí, se une en el surco entre dos subunidades de linfotoxina adyacentes. Esta estructura puede ser representativa de la familia factor de crecimiento neural/receptor de FNT.
INTERLEUCINA-1
La interleucina-1 se produce principalmente por monocitos y macrófagos, pero también por otras células, como células epiteliales y endoteliales. Es un pirógeno endógeno, y muchas de sus funciones son semejantes a las del FNT-alfa. Induce la producción de IL-1 e IL-6 adicional de los macrófagos y también la síntesis de esteroides y liberación de prostaglandina, colagenasa y proteínas de fase aguda. La IL-1 aumenta la expresión de moléculas de superficie en las células endoteliales, causando la adhesión de leucocitos y coagulación, y estimula la producción de quimocinas de macrófagos que a su vez activan neutrófilos. La IL-1 difiere del FNT-alfa en que no produce necrosis de tumores o lesión tisular, aumenta la expresión del complejo principal de histocompatibilidad o, per se, media la reacción de Shwartzman.
La actividad de la IL-1 es resultado de dos polipéptidos, IL-1a e IL-1b, que son producidos por genes separados. La IL-1b es el más abundante. Una proteína de 17 kd, la IL-1b, proviene de la fragmentación de un precursor inactivo de 33 kd por una proteasa de macrófagos. Sin embargo, tanto la molécula IL-1a de 33 kd como la de 17 kd son activas. Los macrófagos producen un antagonista del receptor de IL-1 (IL-1ra) que es estructuralmente semejante a la IL-1 y se une al receptor de IL-1 aunque es inactivo. El IL-1ra, junto con los receptores de IL-1 liberados de las células activadas, inhiben la IL-1 y actúan como reguladores. Estos inhibidores naturales de la IL-1 han sido investigados como agentes potenciales para uso clínico para contrarrestar ciertas proteínas inflamatorias.
INTERLEUCINA-8 Y OTRAS QUIMOCINAS
La IL-8 y otras citocinas quimiotácticas o quimocinas, son importantes para inducir y mantener las reacciones inflamatorias. Diferentes quimocinas pueden unirse al mismo receptor. El peso molecular de las quimocinas varía de 8 a 11 kd, y están clasificadas en dos subfamilias, a y b.
Las moléculas de la subfamilia de quimocinas a se distinguen por su localización en el cromosoma 4 y por su secuencia de aminoácidos, en la que las dos primeras cisteínas están separadas por un aminoácido (Cys-X-Cys). La subfamilia a incluye IL-8, b-tromboglobulina (b-TG), gro-a, gro-b, gro-g y factor plaquetario 4. Suelen ser producidos por monocitos, algunos son producidos por otras células, incluyendo células T, células endoteliales y plaquetas. La IL-8 induce la formación de integrinas de unión a neutrófilos en las células endoteliales, causando acúmulo rápido de neutrófilos en los tejidos. La quimocina gro, también conocida como MIP-2a y MIP-2b, causa también acúmulo de neutrófilos, así como liberacipde enzimas lisosomales que contribuyen a la respuesta inflamatoria local. El factor plaquetario 4 y la b-TG, liberadas de plaquetas agregadas, estimulan a los fibroblastos requeridos para la reparación en sitios de hemorragia o trombosis.
Las moléculas de la subfamilia de quimocinas b se localizan en el cromosoma 17 y tienen un patrón aminoácido cisteína-cisteína (Cys-Cys). Incluyen a RANTES (quimocina expresada y secretada en las células T normales y que regula su activación), al factor quimiotáctico y activador de macrófagos (FQAM), a MIP-2a y a MIP-2b. Las quimocinas de la subfamilia b son producidas en monocitos y células T activadas. RANTES es un atractor potente para células T de memoria, pero no para células T nativas, y también atrae monocitos. El FQAM actúa solo en los monocitos, atrayendo, activándolos, y regulando la expresión de integrinas en su superficie. MIP-1a y MIP-1b atraen solo monocitos.
LINFOTACTINA
La linfotactina, un factor quimiotáctico de linfocitos, fue clonado recientemente de timocitos tempranos (i.e., células pro-T). Este factor puede atraer a los progenitores celulares al timo.55
FACTOR ESTIMULADOR DE COLONIAS DE GRANULOCITOS Y MACROFAGOS
El FEC-GM actúa sobre células de la médula ósea que ya están comprometidas para convertirse en leucocitos. Es producido por células T, macrófagos, fibroblastos y células endoteliales vasculares. Los macrófagnos de la médula ósea cultivados con FEC-GM tienen mayor capacidad para presentar antígenos a células TH1, comparado con los macrófagos de la médula ósea cultivados en factor estimulador de colonias de macrófagos (FEC-M).56 La inyección de macrófagos de médula ósea cultivados con FEC-GM con antígeno de L. major estimula la respuesta de células TH1 en los ratones susceptibles y los hace resistentes al reto con L. major. También se ha encontrado que que el FEC-GM activa macrófagos.57 El FEC-GM es un dímero de 22 kd que no circula sino que actúa en forma local. Se ha usado para contrarrestar la leucopenia en pacientes tratados con quimioterapia.
El FEC-GM es un miembro de la familia de las citocinas hematopoyéticas. Otras citocinas que son principalmente hematopoyéticas incluyen al factor de células tronco, la IL-3, el FEC-M, el FEC-G, la IL-9 y la IL-11.
Papel de los carbohidratos en la regulación de las respuestas de las células T
Estudios recientes sobre la respuesta inmunológica a los parásitos Schistosoma han demostrado que un carbohidrato de superficie, lacto-N-fucopentaosa-III (LNFP-III), puede cambiar una respuesta mediada por células TH1 en una respuesta TH2 mediada principalmente por anticuerpos.58 El LNFP-III estimula las células B convencionales y las células B1 para secretar grandes cantidades de IL-10. La IL-10, al actuar a través de los macrófagos, inhibe las células TH1 y favorece la respuesta TH2.
Esto tiene implicaciones importantes para la imunidad a muchos organismos porque este azúcar, que contiene Lewisx, está presente en otros parásitos y células neoplásicas. El microrganismo o célula neoplásica puede usar el carbohidrato para inhibir la respuesta inmunológica mediada por células protectora del huésped. Este dato puede permitir el desarrollo de una estrategia novedosa para la prevención de la evasión inmunológica por microrganismos y células tumorales, por ejemplo, bloqueando el receptor ß1 usado por el LNFP-III, o el desarrollo de respuestas de anticuerpos más intensas usando LNFP-III en la vacuna. Aún más, recientemente se ha demostrado que la misma familia de oligosacáridos se une a la proteína NKR-P1 de las células NK. Esta interacción es crucial para el reconocimiento de las células blanco por las células NK y para la citotoxicidad.59
Activación de las células B
Cuando un receptor de la célula B es activado por un antígeno soluble, ocurre uno de dos eventos: apoptosis o proliferación y maduración posterior de las células B.60 Las señales para estos procesos se generan dentro de las células por la Iga e Igb, dos proteínas que se asocian con los receptores de la célula B en la misma forma que las proteínas CD3 se asocian con el RCT. La Iga y la Igb reclutan moléculas de transducción de señales en una forma semejante a la del complejo RCT-CD3, específicamente en dos fases de cinasas de tirosina, la primera consistente en p56lyn, p59fyn y p55blk, y la segunda en p72syk. Se reclutan entonces algunas de las mismas proteínas descritas para la activación de la célula T [ver antes, Transducción de señales en Reconocimiento de antígenos por las células T], por ejemplo PI3-cinasa, p95vav, p21ras, p72-74raf, cinasas mek, PLC-b y PLC-g). La vía de activación de las células B es semejante pero no idéntica a la de activación de las células T.
Se sabe relativamente poco sobre la señalización que tiene lugar en las células B cuando presentan un antígeno a una célula T que lo reconoce. Como cuando se activan por antígenos solubles, la interacción célula T-B causa apoptosis o proliferación de las células B. Sin embargo, con la ayuda de la célula T la proliferación no provoca la aparición de diferentes clases de células B.61
CELULAS FOLICULARES EN MANTO
Las células foliculares en manto expresan IgD (IgD+, CD38-) en su superficie y pueden madurar a células plasmáticas que producen solo anticuerpos de IgM. Estas células producen IgM que muestra gran heterogeneidad, pero las células no sufren mutación somática para generar un número aún mayor de moléculas de anticuerpos heterogéneos.
CENTROCITOS
Los procesos que causan mutación somática ocurren en los centrocitos, que son células B IgD-, CD38+ que han sufrido cambio de clase para producir IgG. El paso de maduración de las células B IgD+, CD38- a células B IgD- CD38+ depénde del contacto intercelular con células T antígeno específicas [ver figura 7] y con citocinas. La IL-4 tiene especial importancia para los eventos que causan el cambio de clase.
CELULAS DE MEMORIA
La célula B más madura es la célula de memoria, que no expresa IgD o CD38, pero que se distingue por otros marcadores de superficie y por su localización dentro de los centros germinales. Las células de memoria pueden convertirse en células plasmáticas que producen IgM, IgG, IgA e IgE. Sin embargo, en general la generación de células plasmáticas es independiente de la ayuda de los linfocitos T.
INTERACCION DEL LIGANDO CD40-CD40
Recientemente se ha aclarado la importancia de la interacción entre el ligando CD40 y CD40 (CD40L).62 El CD40L es una glucoproteína tipo 2 que se localiza en la superficie de la célula T relacionada al FNT. La expresión temporal de CD40L en la superficie de las células T es inducida por la unión del RCT con el complejo antígeno-molécula del CPH y por la unión de moléculas B7 con CD28 o CTLA-4. A su vez, la ligadura de CD40 por CD40L causa estimulación del cambio de clase de inmunoglobulinas en las células B. Esto se demostró mejor por el descubrimiento del defecto molecular en una inmunodeficiencia genética denominada síndrome de hiper-IgM. La mutación del gen CD40L es responsable del cambio de clase defectuoso que sucede en este síndrome. Las respuestas de las células B independientes de células T y las respuestas inducidas por el anti-CD40 no se afectan por esta mutación. Por lo tanto, la ayuda de la célula T para activar al linfocito B se realiza principalmente por la vía coestimuladora CD40-CD40L. Los pacientes con síndrome de hiper-IgM también son susceptibles a los patógenos que normalmente son atacados por las células T, produciendo infecciones oportunistas semejantes a las del SIDA, lo que destaca el papel de la coestimulación CD40L en la activación de las células T normales. La ausencia de CD40L tiene un efecto más dramático en el cambio de clase de Ig que la ausencia de células T. Por ello, la suposición lógica es que CD40L podría expresarse en otras células que no son T, pudiendo inducir el cambio de clase en las células B a través de la unión con CD40.
Inmunidad mediada por células
La inmunidad mediada por células consiste en la destrucción de microrganismos invasores como bacterias, virus, hongos y parásitos, la destrucción de células tumorales, el rechazo de injertos tisulares y la lesión a los tejidos en varios estados patológicos, incluyendo la autoinmunidad. Las reacciones inmunes mediadas por células pueden también inducirse por contacto con antígenos, como los encontrados en la hiedra venenosa y muchos medicamentos. Los medicamentos provocan más reacciones mediadas por células cuando se aplican por vía tópica más que sistémica.
La mayoría de las reacciones inmunes mediadas por células incluyen la interacción inicial entre células T sensibilizadas y antigenos en las células presentadoras. Esta reacción puede desencadenar varias vías efectoras, incluyendo activación de células T citotóxicas, estimulación de la producción de citocinas que activan macrófagos y promueven la proliferación de células NK, y producción de anticuerpos incluidos en la CMCDA por células NK y otras células. Aunque las reacciones inmunes mediadas por células diferentes a la CMCDA no requieren la presencia de anticuerpos o complemento, pueden modificarse por estos factores humorales. Los eventos subsecuentes requieren de la cooperación entre los diferentes subtipos de células T. Las reacciones incluidas son controladas por diversas citocinas.
Los mecanismos de inmunidad mediada por células incluyen interacciones entre las células T y macrófagos que pueden ser protectoras (causando destrucción del microrganismo invasor) y dañinas (provocando inflamación y destrucción tisular). En ocasiones, ambas respuestas van de la mano, por ejemplo, en la tuberculosis, tanto la destrucción del bacilo tuberculoso como la formación de cavernas pulmonares son consecuencia de la interacción célula T-macrófago.
CELULAS T CITOTOXICAS
Las células T citotóxicas son células efectoras antígeno-específicas que son importantes para resistir a los agentes infecciosos, en especial a virus existentes en células que no son macrófagos, destrucir tumores y en el rechazo de injertos. La mayoría de las células T citotóxicas con CD8+, y reconocen antígenos presentados por moléculas de clase I del CPH, aunque algunas pocas son T CD4+ y reconocen al antígeno en presencia de moléculas de clase II del CPH. Las células T citotóxicas se desarrollan y sufren selección en el timo, después maduran a células T citotóxicas funcionales en los tejidos. El precursor inmaduro de las células T citotóxicas tiene RCT de superficie y expresa moléculas CD8 pero no es funcional. La células precursora adquiere capacidad lítica después de dos señales. La primera consiste en activación por contacto con el complejo antígeno-molécula del CPH en una CPA. La segunda señal es proporcionada por citocinas, generalmente producida por células T cooperadoras CD4+. Entre las muchas citocinas que se piensa participan en la activación de las células T citotóxicas se incluyen el interferón gama, la IL-2, la Il-4, la Il-7 y la IL-12. Los detalles de la activación de las células T citotóxicas y las moléculas de adhesión que participan se describen en otra parte [ver antes, Procesamiento y presentación del antígeno a las células T]. Las células T citotóxicas desarrollan gránulos que contienen moléculas citotóxicas, incluyendo perforinas (proteínas que producen orificios o poros en la membrana de superficie de la célula), granzima A, granzima B y esterasas de serina. De éstas, las perforinas son las más importantes, como se ha demostrado en ratones en el que se delecionó el gen de la perforina.63 Un segundo mecanismo de destrucción celular incluye la unión del ligando Fas en la célula T citotóxica y la molécula Fas en la célula blanco.64 Este es el único mecanismo disponible para el ratón perforinanulo, y se usa en forma preferencial (aunque no exclusiva) en las células T citotóxicas CD4+. Las células T citotóxicas también producen citocinas, como linfotoxina, interferón gama y FNT-a.
La capacidad de las células T citotóxicas para destruir células blanco infectadas por virus está gobernada por restricciones antigénicas y genéticas. Los linfocitos de ratones que han sido infectados por un virus matarán solo a las células blanco que hayan sido infectados con el mismo virus y porten el mismo antígeno de clase I del CPH. Por lo tanto, para que ocurra la lisis, las células blanco deben portar tanto el mismo antígeno viral como la misma molécula de clase I del CPH que las células que indujeron en forma original a la células T citotóxicas. Los antígenos de clase I del CPH parecen ser los ideales para activar a las células T citotóxicas porque están presentes en todas las células excepto en los eritrocitos. Los estudios han demostrado que las células T citotóxicas pueden ser la principal vía efectora de la inmunidad mediada por células contra ciertos virus en vivo, más que los macrófagos activados por citocinas. Los estudios in vitro han demostrado que si las células blanco se infectan con un virus, pueden ser destruidas después de algunas horas por las células T citotóxicas específicas, antes de que el virus tenga oportunidad de replicarse. Un subgrupo de células T citotóxicas que tienen RCT-gd puede destruir sin restricción del CPH.
CELULAS ASESINAS NATURALES
Las células NK son linfocitos granulares grandes que portan diversos marcadores de superficie celular, incluyendo NK1.1 (un receptor que se pensaba participaba en el reconocimiento de la célula blanco), CD2 y CD16 (un receptor de Fc IgG de baja afinidad, FcgRIII). Las células NK no tienen el marcador complejo RCT-CD3 ni la inmunoglobulina de superficie de las células B. Estas células pueden destruir células infectadas por virus y diversas líneas celulares tumorales in vitro, y se piensa que tienen actividades antivirales y antitumorales. Predominan en la respuesta a la reacción injerto contra huésped después del trasplante de médula ósea. Las células NK no requieren sensibilización previa para volverse citotóxicas, y sus actividades no están restrigidas por el CPH. CD3-s, que funciona en la transducción de señales, se asocia con CD16 en las células NK, y puede detectarse CD3-e en las células NK activadas. En general, se piensa que las células T y las células NK pueden tener un precursor común.
Las células NK no maduran solo en el timo. De hecho, pueden activarse en el ratón deficiente de timo y en el ratón con DICS. La IL-12 es un inductor potente de la proliferación y activación de las células NK, también el interferón gama y la IL-2 activan estas células. La IL-2 aumenta la capacidad tumoricida de las células NK que causa el desarrollo de células LAK. Las células NK pueden también secretar interferón gama, que actúa en forma autócrina para incrementar la actividad de las células NK. El FNT parece contrarrestar el desarrollo de las células NK inducido por IL-12.
Las células NK se fijan a su célula blanco por un receptor en un paso independiente de Ca2+ [ver figura 8]. Sin embargo, una vez que las células NK se activan, requieren iones Ca2+ para la lisis. Esto se logra por orientación de sus gránulos hacia la célula blanco y la liberación de perforinas y enzimas proteolíticas. Destruyen por mecanismos semejantes a los de las células T citotóxicas, como se ha demostrado en el ratón perforinanulo. La destrucción por células NK, al contrario de la destrucción por células T citotóxicas, no es bloqueada por anticuerpos monoclonales anti-CD3 o anti-CD8, pero sí por anticuerpos contra LFA-1.
CITOTOXICIDAD MEDIADA POR CELULAS DEPENDIENTE DE ANTICUERPOS
Las células NK, macrófagos, neutrófilos y eosinófilos se encuentran entre las células que median las reacciones de CMCDA. El anticuerpo, por lo general IgG, se une al receptor Fc en la célula efectora y el sitio de combinación del anticuerpo se une al antígeno en la célula blanco, lo que produce una reacción antígeno específica [ver figura 8]. Por ejemplo, los eosinófilos son eficaces para destruir esquistosómulas cubiertas por anticuerpos, la larva del parásito de Schistosoma. El mecanismo de destrucción incluye la unión de los eosinófilos a la larva a través del anticuerpo unido, seguido de la liberación de proteínas básicas y otras moléculas de los gránulos eosinófilos, causando la formación de poros en la célula blanco. Además de la IgG, la IgE y la IgA pueden mediar también la CMCDA. Los eosinófilos son reclutados a través de la producción de IL-5 por las células T. Los eosinófilos muestran mayor actividad citotóxica después de su activación por citocinas, como el factor estimulador de eosinófilos citotóxicos (producido por macrófagos), el FNT-a, el FEC-GM y la IL-5.
MACROFAGOS ACTIVADOS
Los macrófagos activados son células efectoras importantes en las respuestas inmunes mediadas por células contra varios microrganismos, incluyendo M. tuberculosis, M. leprae, Listeria monocytogenes, Legionella pneumophila, Salmonella, Leishmania y Toxoplasma gondii.65 En esta vía efectora, el macrófago ingiere al microrganismo y después presenta el péptido antigénico procesado en el surco de la molécula de clase II del CPH a la célula T cooperadora CD4+ [ver figura 8]. En la presentación subsecuente al mismo antígeno por el mismo macrófago, las células T cooperadoras sensibilizadas son activadas para producir y secretar varios factores quimiotácticos que atraen más monocitos/macrófagos, granulocitos y linfocitos al sitio de la reacción. Las propiedades de los macrófagos se alteran por FAM, muchos de los cuales son producidos por células T, incluyendo interferón gama (la citocina FAM más importante, así como FEC-GM, FIM, IL-3, IL-4 y FNT-a. El LPS, al actuar con algunas de estas citocinas, aumenta también la activación de los macrófagos [ver figura 9]. La activación de los macrófagos puede ser modulada y disminuida por otras citocinas, incluyendo FTC-b, IL-4 e IL-10, y por prostaglandinas.66
Los FAM inducen la expresión de muchos genes en los macrófagos, causando el fenotipo de activación. Aumenta la fagocitosis y la endocitosis. La mayor expresión de HLA-DR, ICAM-1 y LFA-3 y la inducción de B7 aumenta la presentación del antígeno. La mayor afinidad de los receptores de Fc IgG permite una mejor opsonización de los microrganismos. Los macrófagos activados destruyen microrganismos y células tumorales a través de diversos mediadores, incluyendo la mayor producción de sintetasa de óxido nítrico, lo que causa generación de óxido nítrico y producción de peróxido de hidrógeno y otros radicales reactivos del oxígeno.
La capacidad de los macrófagos activados para destruir microrganismos aumenta en forma inespecífica, esto significa que los macrófagos pueden eliminar no solo al organismo específico que indujo la producción del FAM, sino tambien a otros organismos. Por ejemplo, un macrófago que ha sido activado para destruir L. monocytogenes puede también destruir organismos Salmonella de modo eficaz.
Los macrófagos activados producen muchos mediadores inflamatorios que pueden aumentar la inmunidad atrayendo leucocitos al área, pero también pueden promover la inflamación que causa destrucción tisular. Las sustancias producidas por los macrófagos activados incluyen prostaglandinas, quimocinas, factor activador de plaquetas, factores de coagulación, factor activador del plasminógeno, factor de crecimiento derivado de plaquetas y factor de crecimiento de fibroblastos. Estos factores participan en la inflamación, la coagulación, la proliferación de fibroblastos, la producción de colágena, la fibrosis y la angiogénesis. Las consecuencias patológicas de esta interacción célula T-macrófago es la base de las reacciones de hipersensibilidad tardía.
REACCIONES DE HIPERSENSIBILIDAD TARDIA
Las reacciones de hipersensibilidad tardía están mediadas por células T. Estas reacciones alcanzan su máximo 24 a 48 horas después de la introducción de un antígeno en la piel, a diferencia de la hipersensibilidad alérgica inmediata mediada por IgE, que ocurre en segundos a minutos después de la introducción de un antígeno, y de las reacciones mediadas por complejos inmunes-IgG-complemento (de Arthus), que alcanzan su máximo después de dos a cuatro horas. A diferencia de las reacciones inmediatas y de Arthus que pueden transferirse a los animales humanos por anticuerpos, las reacciones de hipersensibilidad tardía se transfieren solo por células T sensibilizadas.
El prototipo de la reacción de hipersensibilidad tardía es la prueba cutánea a la tuberculina. Cuando se inyecta derivado proteico purificado (PPD) de M. tuberculosis en la piel de una persona que se ha sensibilizado antes por una infección previa, el antígeno es procesado por CPA, incluyendo células endoteliales venulares y se presenta a las células T sensibilizadas con PPD. Esto permite la elaboración de citocinas, la activación de algunos macrófagos y la producción y liberación de más mediadores, como quimocinas, FNT-? e interferón gama. Estos mediadores atraen leucocitos e inducen la expresión secuencial de las moléculas de adhesión vascular ELAM-1, VCAM-1 e ICAM-1, provocando reclutamiento y extravasación de leucocitos en el área [ver figura 10]. Aunque los neutrófilos aparecen pronto en las reacciones de hipersensibilidad tardía, después de 24 horas la mayoría de las células son macrófagos, con algunos linfocitos (la radiación en dosis bajas, que destruye principalmente a los precursores de macrófagos en la médula ósea, disminuye en forma importante el infiltrado celular en la hipersensibilidad tardía) [ver figura 11]. Las lesiones por hipersensibilidad tardía característicamente son firmes o induradas como resultado del depósito de fibrina en la lesión. La hipersensibilidad por contacto, como la observada con la hiedra venenosa o las alergias a medicamentos, es una forma de hipersensibilidad tardía. En algunos casos las lesiones aparecen antes que en la hipersensibilidad tardía del tipo tuberculina, y contienen muchos basófilos. A esta reacción se denomina hipersensibilidad basófila cutánea o de Jones-Mote. Los mecanismos que participan en las reacciones de hipersensibilidad tardía también se observan en procesos granulomatosos y en algunas reacciones autoinmunes.
Bibliografía
DR. JOHN DAVID
DR. COX TERHORST, PH.D.
Interacciones célula-célula
Cuando las células del sistema inmune encuentran un antígeno tiene, lugar una respuesta inmune humoral, celular o ambas. Las respuestas inmunes dependen de linfocitos específicos. La inmunidad humoral es mediada por los linfocitos B, los cuales después de la estimulación proliferan y se diferencían en células plasmáticas productoras de anticuerpos. La inmunidad celular es mediada por los linfocitos T, que al ser activados secretan varias sustancias importantes en la respuesta inmune y que inician diversas reacciones. La primera etapa que conduce a la activación de las células efectoras de ambas divisiones del sistema inmune, consiste en la interacción del antígeno sobre una célula portadora de antígenos (CPA) con el linfocito T cooperador. Los linfocitos B y T se comunican entre sí a través de interacciones que implican receptores (v.gr., interacción entre el receptor de células T [RCT] y el antígeno o la interacción entre varias moléculas de adherencia celular y sus ligandos) y mediante la secreción de varios factores solubles denominados citocinas.1
Procesamiento y presentación del antígeno a las células T
El repertorio de las células T se caracteriza por receptores de superficie celular cuya población muy diversa de regiones variables (V) facilita respuestas muy específicas ante los antígenos. Las células T prototipo ab+ tienen un RCT formado por cadenas a y b (RCT-ab). Estas células responden a antígenos no propios solo cuando los RCT-ab se ponen en contacto con derivados de péptidos antigénicos unidos a moléculas del complejo principal de histocompatibilidad (CPH) localizadas en la superficie celular, un proceso denominado restricción del CPH. El reservorio diverso de las células T se desarrolla conforme las células T inmaduras son sometidas a restricción del CPH dentro del timo durante su desarrollo temprano, y en ese momento son seleccionadas en forma positiva o negativa según su avidez diferencial a antígenos. Si se seleccionan en forma positiva, los linfocitos T con restricción a moléculas del CPH de clase I se convierten en células CD8+, y las que tienen restricción a moléculas de clase II del CPH se convierten en linfocitos CD4+.
Una fracción variable de células T periféricas gdt, tienen un RCT formado por cadenas g y d (RCT-gd), para el que no se conoce hasta el momento un elemento de restricción. Al parecer las células T gdt no requieren presentación del antígeno por las moléculas del CPH.
UNION NORMAL DEL PEPTIDO DENTRO DE LAS MOLECULAS DEL CPH DE CLASE I Y CLASE II
Los productos del CPH son glucoproteínas de superficie celular que unen péptidos antigénicos (i.e., antígenos procesados) dentro de una célula (v.gr., un macrófago) y los transportan a la superficie celular para la interacción con células T [ver adelante, Procesamiento del antígeno]. En el CPH existe un gran polimorfismo (i.e., un gran número de alelos por locus). Sin embargo, dentro de la población humana, cada individuo expresa solo un pequeño número de proteínas diferentes del CPH. Para asegurar una respuesta inmunológica adecuada contra un gran número de antígenos no propios, cada proteína del CPH debe ser capaz de unirse a gran cantidad de péptidos diferentes.
La proteínas de clase I del CPH consisten en una cadena pesada a y una cadena ligera b2-microglobulina (b2-M), mientras que las proteínas de clase II están formadas por cadenas a y b de casi el mismo tamaño [ver la figura 1].2 Las dos moléculas comparten una estructura semeante: dos dominios parecidos a las inmunoglobulinas y un sitio de unión del péptido formado por una hoja de ocho cadenas con plegamiento b y dos regiones a-helicoidales . Mientras que las proteínas de clase I del CPH unen solo péptidos pequeños de longitudes definidas, las proteínas de clase II unen péptidos más grandes, sin aparente restricción por su longitud. Un dato interesante es que ciertos péptidos se unen solo a alelos específicos de las moléculas del CPH de clase I o clase II, por lo que las personas que no tienen el alelo específico no desarrollarán una respuesta inmunológica al péptido asociado.
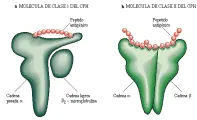
|
| Figura 1 |
| Estructura de las moléculas del CPH |
Con base en estudios de cristalografía y fisicoquímicos se ha propuesto un mecanismo común para la unión de los péptidos a las moléculas de clase I del CPH: los péptidos, generalmente formados por residuos de ocho a 11 aminoácidos de longitud, se unen en el extremo aminoterminal y carboxiterminal al sitio de unión en la molécula de clase I del CPH por medio de una red de uniones hidrógeno a los residuos aminoácidos conservados. Los péptidos más largos se acomodan enrollándose en el centro [ver figura 1a].
La unión de los péptidos a las moléculas de clase II sigue distintos principios.3,4 Estos péptidos tienden a ser mayores y de longitud artribraria. Los residuos aminoácidos conservados forman uniones de hidrógeno en todo el sitio de unión, asegurando la cadena principal del péptido en gran parte de su longitud [ver figura 1b].
Las diferencias en la unión de péptidos entre las proteínas del CPH de clase I y II son causadas por una serie de diferencias estructurales pequeñas dentro de un marco del sitio de unión del CPH relativamente fijo. Estos dos modos de unión probablemente se explican por diferencias fundamentales en el procesamiento de los péptidos que tiene lugar en el retículo endoplásmico (RE) para las moléculas de clase I del CPH y en los endosomas y lisosomas para las moléculas de clase II [ver adelante, Procesamiento del antígeno]. Independientemente de estas diferencias, el área de superficie del péptido central disponible para el reconocimiento directo por un RCT (400 a 500 Å2) es aproximadamente igual para las moléculas de clase I y de clase II.
UNION DEL SUPERANTIGENO CON LAS MOLECULAS DE CLASE II
Un método alternativo de presentación de antígeno incluye a las moléculas de clase II del CPH y a una familia de proteínas que se denominan superantígenos.5 Los superantígenos constituyen una clase de moléculas inmunoestimuladoras asociadas con enfermedades que se unen a moléculas de clase II y estimulan linfocitos T. Los superantígenos han sido implicados en las enfermedades virales, como rabia, síndrome de inmunodeficiencia adquirida (SIDA) y tumores mamarios murinos, y en respuestas de células T a toxinas bacterianas (v.gr., Staphylococcus aureus en el síndrome de choque tóxico y en la intoxicación alimentaria). Aún más, los superantígenos pueden causar enfermedades autoinmunes, como artritis reumatoide. Estudios químicos de las proteínas y cristalográficos han dado evidencias de que los superantígenos se unen a las moléculas de clase II por fuera del sitio convencional de fijación de antígeno. Por ejemplo, el superantígeno bacteriano intacto constituido por la enterotoxina B de S. aureus se une al dominio a1 de la molécula de clase II del CPH humano HLA-DR1. Por lo tanto, se crea un nuevo sitio de fijación al RCT adyacente al sitio de unión convencional del CPH.Aunque los péptidos antigénicos no inhiben la estimulación de las células T por los superantígenos unidos al CPH, los superantígenos pueden bloquear las interacciones convencionales péptido-molécula del CPH con el RCT por obstrucción estérica. Los diferentes alelos de clase II del CPH tienen distintas constantes de unión para superantígenos, causando activación de un repertorio diferente de células T.
PROCESAMIENTO DEL ANTIGENO
Moléculas de clase I del CPH
Las moléculas de clase I del CPH presentan péptidos derivados de proteínas nucleares y citosólicas endógenas degradadas, así como proteínas introducidas por patógenos intracelulares. Para llegar al sitio de unión del péptido de la molécula de clase I del CPH en la luz del RE, los péptidos tienen que cruzar la bicapa lípida del RE. Un sistema de bomba de transporte de péptidos dependiente de trifosfato de adenosina (ATP), que consiste en dos proteínas denominadas TAP1 y TAP2, realiza esta función indispensable. Los requerimientos de esta bomba para pasar el péptido al RE pueden medirse con facilidad porque el ensamble de la cadena pesada a y de la cadena ligera b2M de la molécula de clase I depende de la presencia del péptido. Si no se forma el completo ab2M-péptido, las moléculas de clase I no viajan hasta la superficie celular y son degradadas en la luz del RE. Debido a que las proteínas TAP son polimóficas, aumentan la selectividad de las interacciones péptido-molécula del CPH.
Dos sistemas proteolíticos presentes en todas las células nucleadas se utilizan para atraer a los péptidos hacia la luz del RE para su presentación a las moléculas de clase I. El primero, conjugación ubicua seguida de degradación proteolítica en los proteosomas (subunidades que contienen muchas peptidasas) parece ser critico en la generación de la mayoría de los péptidos presentados [ver figura 2a].6 Sin embargo, el origen preciso de los péptidos no es claro, y pueden originarse en cualquier sitio dentro del citosol. También proteínas no ubicuas y de vida media prolongada pueden ser degradadas en el citosol por los proteosomas. En el segundo mecanismo, antígenos como las proteínas de las cubiertas virales se secretan en la luz del RE, en donde sufren degradación proteolítica por un grupo de proteasas residentes. Como en el caso de los proteosomas, estas proteasas del RE están diseñadas para eliminar de la célula moléculas proteicas sintetizadas en exceso y que no están ensambladas o con constitución inadecuada.
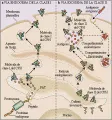
|
| Figura 2 |
| Formación de complejos antígeno-molécula del CPH |
Moléculas de clase II del CPH
Cuando una proteína es introducida a la célula a través de endocitosis, puede ser degradada en los endosomas, lisosomas o en un subcompartimiento endosómico aún no bien definido. Es en estos compartimientos en los que tiene lugar el ensamble entre la molécula de clase II del CPH y el antígeno7,8 [ver figura 2b]. Las proteínas pueden ser procesadas a través de proteólisis antes de la interacción con las moléculas de clase II o, en forma alterna, proteínas antigénicas parcialmente desdobladas pueden unirse a las moléculas de clase II del CPH antes de sufrir degradación proteolítica. Esto puede explicar porqué la longitud de los péptidos unidos a las moléculas de clase II del CPH puede ser muy variable. Después de que el péptido se ha unido a la molécula de clase II del CPH, el endosoma que contiene este complejo migra a la superficie celular y se fusiona con la membrana plasmática.
Aunque la degradación semejante al proteosoma no participa en el procesamiento de los péptidos que se unen a las moléculas de clase II del CPH, dos de los elementos reguladores claves ayudan a controlar las interacciones péptido-molécula de clase II: la cadena invariante y el gen HLA-DM. La cadena invariante, o Ii, bloquea el surco del péptido durante el ensambre de las subunidades a y b de las moléculas de clase II del CPH, evitando así la unión de péptidos procesados por el RE. Es degradada o eliminada del complejo en el endosoma, lo que permite al péptido ocupar el sitio de unión. Se ha demostrado que el gen HLA-DM codifica una función accesoria en el ensamble del complejo molécula clase II del CPH-péptido.9-11 El análisis de las formas mutantes de HLA-DM indican que este producto genético (una proteína semejante a la cadena ? de clase II del CPH) es indispensable en la formación del complejo.
Otras moléculas
Un tercer grupo de moléculas presentadoras de antígeno son las proteínas no polimóficas parecidas a las de clase I asociadas a la b2M. Estas incluyen los productos de los genes no polimóficos CD1 (Cd1a, CD1b, CD1c y CD1e) que se localizan en el cromosoma 1, en lugar de en el cromosoma 6 como las del CPH. Se ha demostrado que CD1b presenta lípidos del ácido micólico de Mycobacterium tuberculosis a células T ab+.12 Este ejemplo de reconocimiento de antígeno no proteico microbiano sugiere que las células T ab+ reconocen un rango de antígenos más amplio de lo que antes se pensaba.
CELULAS PRESENTADORAS DE ANTIGENO PROFESIONALES
Aunque las moléculas de clase I del CPH se expresan en la superficie de todas las células eucariotas, las moléculas de clase II tienen una distribución tisular más restringida. De hecho, ciertas células, incluyendo linfocitos B, células dendríticas, macrófagos, células de Langerhans y células endoteliales, se denominan células presentadoras de antígeno profesionales porque presentan péptidos antigénicos junto con moléculas de clase II del CPH con más eficacia que otras CPA. Esta eficacia se atribuye principalmente a su capacidad para procesar antígenos endocitados. Las CPA profesionales también interactúan con más eficacia con las células T porque tienen varios marcadores de superficie celular, como B7, que se fija a contraestructuras de la superficie de los linfocitos T. Por lo tanto, con frecuencia se realiza una distinción operativa entre las CPA profesionales y no profesionales.
PRESENTACION DEL ANTIGENO A LAS CELULAS T
La detección de complejos molécula del CPH-péptido por las células T ab+ es un proceso dinámico que incluye interacciones intercelulares complejas antes de que tenga lugar la activación del linfocito T. El reconocimiento del antígeno por la célula T sucede en dos fases distintas: (1) un paso de adhesión no específica antes del reconocimiento antigénico, seguido por (2) una interacción específica entre el RCT y el complejo antígeno-molécula del CPH. Este proceso incorpora dos propiedades fundamentales del linfocito T: su capacidad para migrar por todo el organismo y adherirse a muchos tipos de células y su gran especificidad por antígenos particulares.
Las CPA pueden procesar en forma simultánea muchos antígenos proteicos y pueden expresar en su superficie un gran número de complejos diferentes antígeno-molécula del CPH. Solo un número pequeño de estos complejos puede ser reconocido por un determinado linfocito T ab+. A través de la adhesión intercelular, las células T buscan a las CPA potenciales. La adhesión se aborta en ausencia de reconocimiento específico del antígeno por el RCT y se intensifica cuando se logra el contacto correcto RCT-molécula del CPH-péptido. Al adherirse, los complejos RCT-CD3 se agregan en la superficie de membrana de la célula T y se unen a complejos molécula CPH-péptido que se han agregado en la superficie de la CPA.
El orden de los eventos en la búsqueda célula T-CPA se estudió por primera vez en el caso de la función efectora de las células T citotóxicas.13 La fase de adhesión célula T citotóxica-célula blanco dependiente de Mg+2 es seguida de una fase dependiente de Ca+2 que causa la entrega de químicos citotóxicos a la célula blanco. La célula T citotóxica se disocia entonces de la célula blanco, y ocurre muerte de la célula blanco en ausencia de la célula T citotóxica, que recicla para atacar a otra célula blanco. Si la célula T citotóxica que se adhiere a la célula no porta la combinación molécula del CPH-péptido correcta, no se liberan químicos citotóxicos y las células se disocian con más rapidez.
La adhesión de células citotóxicas T a células blanco causa activación de las células T citotóxicas y rearreglo extenso de las estructuras en el citoplasma de la célula T citotóxica. El centro organizador de microtúbulos y el complejo de Golgi se reorientan hacia el sitio de contacto de la célula T citotóxica y la célula blanco. También se acumulan componentes vesiculares en la proximidad del sitio de contacto.
Los anticuerpos monoclonales y las clonas de células T antígeno-específicas fueron cruciales para demostrar que el contacto inicial entre una célula T citotóxica y una célula blanco no requiere de la interacción entre el RCT y su antígeno blanco. Los anticuerpos dirigidos contra las moléculas de adhesión antígeno-1 asociado a la función leucocitaria (LFA-1) y CD2 inhiben las adhesiones tanto específicas como no específicas entre la célula citotóxica y la célula blanco. En forma semejante, anticuerpos monoclonales dirigidos contra las moléculas de adhesión LFA-3 y molécula de adhesión intercelular ICAM-1 bloquean la adhesión a nivel de la célula blanco. Por el contrario, ensayos en agar suave muestran que los rearreglos anticlonotípicos RCT y anti-CD3 no inhiben la adhesión pero sí bloquean la fase lítica. Los anticuerpos anti-CD4 y anti-CD8 inhiben tanto la adhesión como la fase lítica.
Las interacciones entre las células T y las CPA tienen el mismo patrón general. Experimentos que emplearon complejos solubles de molécula de clase II del CPH-péptido sugieren que la constante de unión entre el RCT y este complejo es baja.14 Es probable que solo un pequeño número de complejos molécula CPH-péptido se pongan en contacto inicial con el RCT específico en un área de contacto intercelular establecido por la adhesión. En consecuencia, los complejos RCT-CD3 y las moléculas del CPH que portan el péptido correcto migran hacia al región de contacto célula T-CPA. Esto establece una densidad local muy alta de compleos RCT-CD3, que promueve la unión de antígenos y da lugar a activación del linfocito T. La activación causa reclutamiento de las moléculas de transducción intracelular y un aumento temporal en la adhesión. La activación causa relutamiento de moléculas de transducción intracelular y un aumento temporal en la adhesión. La menor regulación de las moléculas de adhesión permite que la célula se libere. En ausencia de reconocimiento del antígeno, los complejos RCT-CD3 son incapaces de acumularse y ocurre liberación inmediata.
Varias observaciones recientes apoyan este modelo. Los complejos RCT-CD3 tienen propensión a formar agregados en la superficie de la célula T, mientras que las moléculas de clase II del CPH forman dímeros en la superficie de la CPA. Se ha demostrado que se encuentran correceptores CD8 en la superficie de linfocitos T humanos y timocitos como dímeros y multímeros. Por lo tanto, es probable que la interacción receptor-ligando en la interfase del linfocito T con la CPA cause agregación del receptor.
MOLECULAS DE ADHESION
Las principales moléculas de adhesión en las células T son CD2 y LFA-1, que está formada por una cadena alfa (o CD11a) y una cadena b (o CD18).15,16 El ligando natural en la superficie celular para CD2 es LFA-3, o CD58, mientras que el ligando de LFA-1 es ICAM-1 o CD54. Estos ligandos se expresan en todas las CPA potenciales [ver tabla 1]. CD4 y CD8 constituyen una categoría diferente en el paso de adhesión porque, al igual que el RCT, sus ligandos naturales son las glucoproteínas de superficie de clase II y clase I respectivamente del CPH. Las interacciones directas entre CD8 y las moléculas de clase I y entre CD4 y las de clase II tienen un papel crucial en la selección positiva durante la maduración intratímica de los linfocitos T [ver figura 3]. El fenotipo de una célula T determina su función efectora: las células T que expresan CD4 reconocen a las moléculas de clase II del CPH, mientras que las células T que expresan CD8 tienen afinidad por moléculas de clase I. Las moléculas de adhesión celular de los linfocitos T (v.gr., CD2, CD4, CD8 y LFA-1) tienen también una función en la transducción de señales y en la activación de la célula T, que causa una mayor avidez en las interacciones RCT-ligando. A su vez, la transducción de la señal del RCT aumenta la afinidad entre las moléculas de adhesión LFA-1 e ICAM-1.
|
|
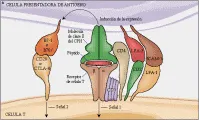
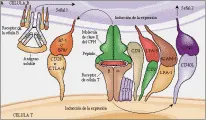
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
CD28, un homodímero asociado a disulfuros formado por dos cadenas glucoproteicas de 44kd, es miembro de la superfamilia de genes de inmunoglobulina, específicamente es miembro de la subfamilia de moléculas que tienen un solo dominio V. CD28 se expresa en todas las células T CD4+ y en la mayoría de las CD8+. Una estructura complementaria, denominada B7-1 (o también CD80) se expresa en menor densidad en células B en reposo y puede facilitar la activación de los linfocitos B. B7-1 puede ser inducido en la superficie de las CPA profesionales, como los macrófagos y las células dendríticas.
Otro antígeno de las células T, CTLA-1, se une a B7-1 con mayor actividad y es homólogo a CD28. Se ha descrito una segunda estructura complementaria para CD28 y CTLA-4 en ratones (B7-2 o CD86) y en humanos (B70 o CD86). El perfil de expresión de B7-2 es semejante al de B7-1, con la importante excepción de que B7-2 se expresa en células dendríticas y monocitos en reposo. Se ha demostrado que ratones con deficiencia de CD28 tienen menores respuestas inmunológicas dependientes de células T, lo que demuestra la importancia de este ligando en la activación de las células T.
Transducción de señales en las células T al reconocer al antígeno
En respuesta al reconocimiento del antígeno, las células T en reposo sufren una serie de eventos complejos conocidos como activación.16-18 La señal del RCT no es suficiente per se para causar que las células T en reposo secreten cantidades máximas de interleucina-1 (IL-2), una citocina necesaria para la proliferación de las células T. Por ejemplo, las células T purificadas no producen IL-2 en respuesta a los anticuerpos anti-RCT en ausencia de células accesorias viables. Además, los linfocitos B activados y en reposo tienen capacidad muy diferente para estimular a las células T incluso si el número de complejos molécula CPH-péptido es idéntico. Estas observaciones indican que las CPA hacen más que presentar al antígeno, también proporcionan señales coestimuladoras. Las moléculas de adhesión de las células T CD4, CD8, y CD28 y CTLA-4 también proporcionan señales coestimuladoras importantes.
SEÑALES DEL RECEPTOR DE LA CELULA T
Se han identificado varios eventos bioquímicos que ocurren inmediatamente después del reconocimiento del antígeno por la célula T. El primero parece ser el reclutamiento de cinasas de tirosina-proteína por los elementos CD3-e y CD3-z del complejo RCT-CD3 y quizá por los elementos CD3-g y CD3-d. Esto causa fosforilación tirosina de las colas citoplásmicas de CD3-s y CD3-e y, en menor grado, de CD3-g y CD3-d. Dos tirosina cinasas principales, p56lck y p59fyn (ambas relacionadas con la familia del proto-oncogén src), participan en la tirosina fosforilación de CD3. El nivel de fosforilación de CD3-s depende de la naturaleza del complejo molécula CPH-péptido reconocido por el RCT-ab.
Cuando se une al complejo RCT-CD3, la p59fyn fosforila y activa a la enzima celular fosfolipasa C-g (PLC-g), que induce la liberación de Ca2+ de las reservas intracelulares al generar inositol 1,4,5-trifosfato (IP3). Esta liberación de Ca2+ es la característica de la activación celular en muchos sistemas, incluyendo las células T y B. Durante el reconocimiento del antígeno, la fosforilación de las colas citoplásmicas CD3-e y CD3-s causan reclutamiento de dos tirosina cinasas diferentes, p72syk y ZAP-70, al complejo RCT-CD3. Las proteínas blanco precisas para p72syk y ZAP-70 se desconocen, pero pueden incluir moléculas inhibitorias de transducción de señales, como p95vav y p72-74raf [ver figura 4].
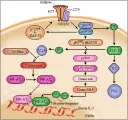
|
| Figura 4 |
| Vías de señales después de la estimulación del RCT |
El ciclo de fosforilación-defosforilación de tirosina es necesario para la activación de la célula T. Las cinasas de tirosina son vitales en estas reacciones. Se ha demostrado que los inhibidores de estas enzimas bloquean las fases tempranas de activación de las células T, mientras que la depleción de la proteína cinasa C (PCC) no lo hace. También es importante la presencia de CD45, una fosfatasa tirosina unida a la membrana. La ausencia de CD45 en las células T inhibe la vía de activación en varios grados, la reconstitución de CD45 por transfección con ADN complementario (ADNc) de CD45 restablece su actividad total. Se sabe que la cinasa carboxi terminal src (csk) inhibe la producción de IL-2 inducida por el RCT en modelos experimentales: csk fosforila los sitios reguladores inhibitorios en las cinasas relacionadas a src p59fyn y p56lck, inactivando estas enzimas. La importancia de la fosforilación de tirosina en la activación de la célula T ha sido demostrada por estudios de ratones mutantes en los que se han alterado los genes fyn, lck o CD45. En otro estudio, los pacientes que no tenían la cinasa de tirosina ZAP-70 fueron incapaces de reclutar células T CD4+. Sorprendentemente, la ausencia de ZAP-70 causó también ausencia de células T CD8+.19,20
La activación de la PLC-? causa la liberación de diacilglicerol, que a su vez activa a la PCC. La PCC inicia la cascada de otras cinasas de serina-treonina, incluyendo p72-74raf, mek y MAP. La molécula central en la transducción de señales, ras, parece participar en eventos más tardíos, después de la activación de PCC. Tanto la vía ras como las vías de cinasa de serina-treonina están acopladas, porque activan genes tempranos como jun y fos, que están implicados en la activación del gen de la IL-2 [ver figura 4]. El Ca2+ generado por el IP3 activa otra cascada, la vía de la calcineurina. La calcineurina, una fosfatasa de serina-treonina, es una enzima dependiente de calcio y calmodulina que participa en la inducción del factor de transcripciòn NF-AT, que se requiere para la transcripción de IL-2. La vía dela calcineurina es bloqueada por la ciclosporina, que se fija a la ciclofilina, inhibiendo la actividad de la calcineurina.
Otra enzima importante que participa en el metabolismo lípido es la fosfatidilinositol-3 cinasa (PI3-cinasa), una enzima clave en las vías de señales mitogénicas de las cinasas de tirosina tanto del receptor como no de receptor. La PI3-cinasa tiene dos propiedades características: fosforilación de la molécula de inositol por el fosfatidilinositol para producir PI3P y asociaciones específicas con muchas cinasas de proteína activadas. La activación del complejo RCT-CD3 aumenta la actividad de la PI3-cinasa, que se asocia con una cadena fosforilada CD3-s. Aún no se ha determinado si la PI3-cinasa se une en forma directa al CD3-s activado o a través de otras cinasas, como las cinasas de la familia src (v.gr., p56lck y p59fyn) y las cinasas p72syk y ZAP-70.
SEÑALES DE MOLECULAS COESTIMULADORAS
La activación aberrante del RCT puede iniciar respuestas inmunológicas muy dañinas, como las observadas en las enfermedades autoinmunes y otras patologías. Las moléculas coestimuladoras tienen un papel muy importante en la coordinación y regulación cinética de la activación de las células T. En algunos casos las moléculas coestimuladoras de los linfocitos T y sus ligandos en las CPA son inducidas por citocinas y transducción de señales después de la interacción intercelular inicial. Estas moléculas pertenecen a diferentes familias de receptores, incluyendo la superfamilia de las inmunoglobulinas, las superfamilias del factor de necrosis tumoral (FNT) y de los receptores de FNT, las integrinas y las selectivas. Las moléculas coestimuladoras de los linfocitos T y otras moléculas cuya unión puede, en algunos casos, inhibir la activación celular son CD2, CD4, CD5, CD7, CD8, CD27, CD28 y CTLA-4, gp39 (CD40L), CD45, LFA-1 (complejo CD11-CD18), ICAM-1 (CD54) y Fas (CD95) [ver tabla 1].
CD28 y CTLA-1 proporcionan la señal secundaria más importante, iniciando una vía de transducción de señales que es diferente e independiente de la vía mediada por el complejo RCT-CD3. CD28 utiliza una cinasa de tirosina recién descubierta, p72.tsk 21 A diferencia de la señal del complejo RCT-CD3, la señal enviada por CD28 no es bloqueada por ciclosporina. Por lo tanto, la vía de transducción de señales de CD28 puede evitar a la fosfatasa dependiente de Ca2+ calcineurina, lo que apoya el concepto de que esta vía es independiente de Ca2+. Además de tener un papel en la activación de la célula T, la activación de CTLA-4 por B7 provoca la inhibición de la fase tardía de la activación de la célula T. CD28 se expresa en forma constitutiva en la superficie de los linfocitos T, pero CTLA-4, un homólogo de CD28, se expresa solo en los linfocitos T activados. Ni B7-1 ni B7-2 se expresa en forma constitutiva en la superficie de los linfocitos B. Un aspecto que aumenta aún más la complejidad es el hecho de que B7-1 y B7-2 son, per se, capaces de realizar transducción de señales en las células B.
La molécula coestimuladora CD27 se induce al activarse la célula T y actúa en forma sinérgica con la activación asociada al RCT para inducir la proliferación de linfocitos T. CD27 (en los linfocitos T) es un miembro de la familia del receptor del FNT, mientras que CD70, la estructura correspondiente en linfocitos B activados, es codificada por el gen de la superfamilia del FNT.
Otro miembro de la familia del receptor de FNT, Fas (CD95) ha sido implicado en señales tanto positivas como negativas a través de la interacción con el ligando de Fas. La unión del antígeno Fas en las células T típicamente causa muerte celular programada (i.e., apoptosis). Sin embargo, se ha demostrado que la unión con un anticuerpo monoclonal anti-Fas permite que Fas funcione como una molécula coestimuladora para la activación del RCT-CD3. Por lo tanto, una sola molécula puede originar diferentes señales en diferentes estadios del desarrollo del linfocito T.
PAPEL DE CD4 Y DE CD8
Las moléculas CD4 y CD8 son correceptores del RCT que interactúan con las moléculas del CPH en la superficie de las CPA. CD4 es un monómero de 55 kd que consiste en cuatro dominios semejantes a los de las inmunoglobulinas, mientras que CD8 está formado por dos cadenas, a y b, que son codificadas por genes diferentes. CD8-b se expresa solo en unión con CD8-a, pero CD8-a puede presentarse como homodímero. Se ha propuesto que el sitio de unión de CD8 es el dominio alfa3 de la molécula de clase I del CPH. Es probable que el contacto entre CD4 y la molécula de clase II del CPH incluya los primeros tres dominios semejantes a inmunoglobulinas de CD4.
El hecho de que CD4 y CD8 se asocien con la cinasa de tirosina citoplásma p56lck sugiere que participan en la transducción de señales. Se requieren dos residuos de cisteínas en regiones claves de CD4 y CD8-a para la asociación con p56lck. Los anticuerpos anti-CD4 pueden inhibir las respuestas de linfocitos T en ausencia de interacción del RCT con antígenos del CPH, lo que indica que CD4 puede trasmitir también una señal denominada negativa.
La interacción de las moléculas CD4 y CD8 con las moléculas del CPH es indispensable para generar a los linfocitos T CD4+ y CD8+. El cambio de timocitos CD4+, CD8+ (dobles positivos) a CD4+ o CD8+ (positivos únicos) requiere de selección positiva en el timo, que depende de la interacción entre el complejo RCT-CD3 con los antígenos del CPH. El ratón clase Inulo del CPH (obtenido por interrupción dirigida del gen que codifica a la b2M o del gen que codifica a TAP1) tiene un número mínimo de timocitos CD8+ comparado con el ratón normal. En forma semejante, el ratón clase IInulo del CPH no tiene timocitos CD4+. El ratón CD8nulo no posee células T citotóxicas, mientras que el ratón CD4nulo no tiene células T cooperadoras. La falta de células T CD4+ y CD8+ en el ratón CD4nulo y CD8nulo, respectivamente, no fue compensada por el subgrupo celular recíproco. Además, el número de células CD4-, CD8- aumentó, demostrando que se requiere de las moléculas CD4 y CD8 para generar células T CD4+ y CD8+ funcionales.
Los mecanismos de transducción de señales que participan en la selección positiva no se han definido aún. Debido a que también se encontró que la cinasa de tirosina p56lck se asocia con CD4 y CD8 en el timo, existe la posibilidad de que esté involucrada en la selección positiva. Sin embargo, su asociación con CD4 y CD8 no es indispensable, ya que pueden desarrollarse células T CD4+ en ratones con CD4 que no tiene cola citoplásmica, lo que le hace incapaz de asociarse con p56lck. En forma semejante, Loh y colaboradores demostraron en 1993 que la asociación de p56lck con CD8 no era necesaria para el desarrollo de células T CD8+.22
Citocinas y receptores de citocinas
Las citocinas, un grupo diverso de proteínas producidas por varios tipos celulares, son críticas en la regulación de las respuestas inmunológicas.23 También son importantes en la diferenciación de los sistemas celulares. En general, las citocinas no se almacenan en las células, sino que se sintetizan por el estímulo apropiado. El ARN mensajero (ARNm) resultante tiene vida media corta. En algunos casos la citocina activa es liberada a partir de un precursor inactivo por proteólisis. Después de su secreción, las citocinas tienen acción local, pueden actuar sobre muy diferentes células, y distintas citocinas pueden tener actividades semejantes. Los receptores de citocinas comparten también cadenas proteicas comunes.24 Por ejemplo, los receptores de citocinas IL-2R, IL-4R, IL-7R, IL-9R, IL-13R e IL-15R comparten cadenas g comunes.25,26 La cadena polipeptídica gp130 es compartida por los receptores de la IL-6, el factor inhibidor de leucemia (FIL) y el factor neurotrópico ciliar (CNTF). Los receptores para IL-3, IL-5 y el factor estimulador de colonias de granulocitos y macrófagos (FEC-GM) comparten una cadena b en común [ver tabla 2]. La mayoría de las citocinas forman una red que regula la activación de las células y la producción de otras citocinas.
|
|
|
Las citocinas tienen un papel importante en el desarrollo y regulación de las células T, De hecho, los subgrupos TH1 y TH2 se definen por las citocinas que producen.
Los linfocitos TH1 se desarrollan bajo la influencia de la IL-2 y el interferón gama. Producen interferón gama, IL-2 y FNT-alfa y participan principalmente en las reacciones de inmunidad mediada por células e hipersensibilidad. El interferón gama es el factor activador más importante para los macrófagos.
Las células TH2 se desarrollan por influencia de IL-4 y participan sobre todo en reacciones alérgicas, producción de anticuerpos y cambio de clase de anticuerpo (en especial de IgE a IgA). Estas células producen IL-4, IL-5, IL-6 e IL-10. La IL-10 inhibe la activación de los macrófagos, lo mismo que la IL-4. Cada uno de estos dos subgrupos de linfocitos T cooperadores inhibe el desarrollo y función del otro. El interferón gama producido por las células TH1 inhibe el desarrollo y función de las células TH2. Por el contrario, la IL-4 y la IL-10 producidas por las células TH2 inhibe el desarrollo y función de los TH1 [ver figura 5]. La IL-12 aumenta la función de las células TH1 porque es un potente factor de crecimiento para las células asesinas naturales (NK), que también producen interferón gama.
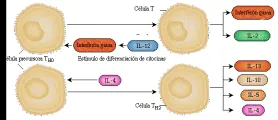
|
| Figura 5 |
| Diferenciación de células T CD4 |
El hecho de que las células TH2 inhiban las respuestas inmunológicas mediadas por TH1 y que las TH1 inhiban las respuestas mediadas por anticuerpos de las TH2 puede atribuirse a que ambos subgrupos inducen también inflamación, que debe controlarse [ver adelante, Reacciones de hipersensibilidad tardía].
INTERLEUCINA-2
La IL-2 es producida por las células T después de la estimulación por un antígeno o mitógeno, y es el principal factor de crecimiento para las células T CD4+ y CD8+. También influye en el desarrollo de los linfocitos B. La IL-2 estimula el crecimiento y la función citolítica de células NK específicas conocidas como células activadas por linfocinas (LAK). Dos retrovirus, el virus linfotrópico de células T humanas tipo I (HTLV-I) y el HTLV-II, producen una proteína transactivadora que estimula la expresión de los genes que codifican tanto para la IL-2 como para su receptor. Los anticuerpos monoclonales contra el receptor de la IL-2 tienen un papel potencialmente terapéutico en la anemia aplástica y la leucemia causada por HTLV-I.
La IL-2 es una glucoproteína de 15 a 17 kd (heterogénea por glucosilación diferente). Es miembro de la familia de citocinas hematopoyéticas y, como todos los miembros, está compuesta de cuatro segmentos de alfa-hélice organizados en dos haces paralelos. La IL-2, la IL-4 y el FEC-GM tienen una estructura muy semejante, y se unen a receptores similares.27 El gen que codifica para la IL-2 se localiza en el cromosoma humano 4. La actividad de la IL-2 no es especie específica.
Se ha demostrado la importancia de la IL-2 por estudios de ratones mutantes con gen de la IL-2 alterado (ratón IL-2nulo): la mitad de estos ratones mueren por anemia severa causada por autoanticuerpos dirigidos contra eritrocitos.28 Los que no mueren por anemia lo hacen por colitis ulcerativa a menos que se mantengan en un ambiente libre de gérmenes. La IL-2 puede ser importante en la eliminación de células autorreactivas.29 Los ratones en los que se alteró el gen que gobierna la cadena ? del receptor de IL-2 (ratones IL-2Rbnulo) murieron por anemia severa a menor edad que los ratones IL-2nulo. La diferencia entre los dos ratones mutantes radica en el hecho de que la IL-15 aún puede funcionar en el IL-2nulo, pero no en el IL-2Rbnulo, porque la IL-2Rb también es parte del receptor de la IL-15.
INTERLEUCINA-15
La IL-15, recién aislada del sobrenadante de cultivos de líneas celulares de epitelio de riñón de simios, apoya la proliferación de una línea celular dependiente de IL-2.30 Estimula a las células que usan las cadenas b y g del receptor de la IL-2, pero no a las que usan la cadena a. Los anticuerpos contra IL-2R bloquearon la actividad de la IL-15. Esta citocina es una proteína de 14 a 15 kd que no tiene homología en su secuencia de aminoácidos con ninguna otra citocina. Un factor semejante de 14kd, denominado como IL-T, también tiene propiedades semejantes a la IL-2 que estimulan la proliferación de las células T e inducen la formación de células LAK. Se ha encontrado que la IL-T no es bloqueada por anticuerpos contra IL-2 o IL-2Ra, pero sí por anticuerpos contra IL-2Rb e IL-2Rg. Las consecuencias de que la cadena polipéptida gama es compartida en los receptores para IL-2, IL-4, IL-7, IL-9, IL-13 e IL-15 se han demostrado ampliamente en pacientes con inmunodeficiencia combinada severa (IDCS) ligada al cromosoma X, que es causada por falta de la cadena polipeptídica gama.
INTERLEUCINA-4
La IL-4, producida principalmente por linfocitos T, es crucial en el desarrollo del subtipo de células TH2 a partir de las células precursoras TH0.23 Las células TH2, a su vez, producen IL-4 adicional, así como IL-5, IL-6, IL-10 e IL-13. Estas células participan en la ayuda a linfocitos B, y la IL-4 es la principal citocina que causa el cambio en la producción de anticuerpos a IgE, y en menor grado a IgG1, en las células B. La IL-4 es un factor de crecimiento para células cebadas y estimula a la molécula de adhesión de células vasculares-1 (VCAM-1) sobre las células endoteliales, aumentando su capacidad para fijar eosinófilos, linfocitos y monocitos. Las células cebadas activadas y los basófilos producen IL-4 adicional. Las células TH2 son importantes para mediar las reacciones alérgicas, así como la inmunidad a nemátodos, en la que los anticuerpos tienen un papel muy importante. Por otro lado, la IL-4 inhibe el desarrollo del subgrupo TH1 y los efectos activadores sobre macrófagos del interferón gama, incluyendo la actividad microbicida y la producción de IL-1 y de óxido nítrico. Por lo tanto, la IL-4 inhibe la inmunidad protectora contra organismos intracelulares y suprime los procesos inflamatorios inducidos por las células TH1.
La IL-4 es una glucoproteína de 20 kd miembro de la familia de citocinas hematopoyéticas. Se codifica en el cromosoma humano 5, cerca del gen de la IL-5. La actividad de la IL-4 es muy específica de especie. El ratón IL-4 nulo tiene alteración en el cambio de isotipo de inmunoglobulinas y células TH2 parcialmente disfuncionantes.31,32 La IL-13 tiene la capacidad de realizar algunas de las funciones de la IL-4.33
INTERLEUCINA-13
La IL-13, producida por las células T CD4+ y CD8+ activadas, comparte muchas de las funciones de la IL-4.33 Actúa sobre los macrófagos para suprimir varias citocinas proinflamatorias, quimocinas (i.e, citocinas quimiotácticas) y factores de crecimiento hematopoyético (incluyendo IL-1, IL-8, FNT-a, proteína inflamatoria de macrófagos-1a [PIM-1a] y FEC-GM), inhibe la citotoxicidad mediada por células dependiente de anticuerpos (CMCDA) al disminuir el número de receptores Fc, e inhibe la producción de óxido nítrico en macrófagos murinos, y con ello la capacidad de los macrófagos para destruir parásitos intracelulares, como Leishmania. Sin embargo, la IL-13 aumenta la presentación de moléculas clase II del CPH y antígenos. A diferencia de la IL-4, la IL-13 actúa sobre los linfocitos B humanos para cambiar la producción de anticuerpos a IgE e IgG4 (pero no lo hace en linfocitos B murinos). Aunque diferentes, los receptores IL-4R e IL-13R comparten algunas características. El cambio de un aminoácido en la IL-4 provoca una molécula que se ha denominado hIL-4.Y124D, que no tiene la actividad de la IL-4 pero aún se une al receptor e inhibe la acción tanto de IL-4 como de IL-13. La capacidad de esta IL-4 mutada para inhibir la síntesis de IgE por la IL-4 y la IL-13 puede ser útil en algunas estrategias de tratamiento.
La IL-13 es una proteína de 10 kd que comparte alrededor del 30 porciento de aminoácidos homólogos con la IL-4. Se transcribe antes que la IL-4 en los linfocitos T activados y durante un periodo más prolongado.
INTERFERON GAMA
El interferón gama, también llamado tipo II o interferón inmune, es producido por las células T y NK. Es crucial en el desarrollo de los linfocitos TH0 a TH1 e inhibe el desarrollo de células TH2. Es el principal factor activador de macrófagos (FAM) y también es una potente citocina en la inmunidad mediada por células. Otros FAM incluyen al FEC-GM y al factor inhibidor de la migración (FIM). La IL-1 y el FNT-a tienen también leve actividad de FAM. La IL-12 estimula a las células NK para producir mayores cantidades de interferón gama, aumentando las reacciones dependientes de esta citocina. El interferón gama aumenta la expresión de moléculas de clase I y clase II del CPH, aumentando la presentación de antígenos y otras reacciones cognoscitivas. Per se y por los mayores efectos del FNT-a, el interferón gama causa la expresión de moléculas de adhesión en la superficie de las células del endotelio vascular., provocando adhesión y extravasación de linfocitos T. El interferón gama se ha empleado con éxito para tratar enfermedades granulomatosas crónicas y leishmaniasis visceral resistente al tratamiento.34
El interferón gama está constituido por dos subunidades de 21 a 24 kd, es heterogéneo por diferencias en la glucosilación. El ratón interferón gamanulo y el ratón con receptor de interferón gamanulo tienen destrucción defectuosa de microrganismos por macrófagos, menor expresión de moléculas de clase II del CPH en los macrófagos y menor producción de óxido nítrico, función deficiente de las células NK y menor concentración sérica de IgG2 e IgG3.
FACTOR DE TRANSFORMACION DE CRECIMIENTO-b
El factor de transformación de crecimiento-b (FTC-b) es producido por diferentes células, incluyendo plaquetas, linfocitos, macrófagos activados y células de la placenta. Está unido en forma no covalente a una proteína asociada de latencia (PAL) de 75 kd, de la cual se libera por acción de una proteasa para adoptar su forma activa. El FTC-b permite a las células normales crecer en agar suave, una característica que suele observarse en células malignas. El FTC-b es una citocina muy pleiotrópica, estimula la producción de proteínas de la matriz extracelular, como la colágena. También cambia a producción de IgA en los linfocitos B (aumentando así las reacciones inmunológicas mucosas) e inhibe la producción de IgM, IgG1 e IgG2. Además, el FTC-b inhibe la activación de macrófagos, la proliferación de linfocitos inducida por mitógenos y la maduración de células T citotóxicas, controlando los efectos de muchas citocinas. De hecho, las reacciones inflamatorias en los ratones FTC-bnulo se controlan muy mal.
El FTC-b está constituido por tres péptidos homodiméricos de 25 kd (FTC-b1, FTC-b2 y FTC-b3) y cada isoforma es codificada por genes diferentes mapeados en distintos cromosomas.
INTERLEUCINA-10
La IL-10 es producida por linfocitos T CD4+ y CD8+, linfocitos B, macrófagos, células cebadas y queratinocitos.36 Aunque casi siempre se asocia con actividad de células TH2, también puede producirse por células TH1.37,38 La IL-10 suprime las respuestas de linfocinas al disminuir la producción de citocinas por los macrófagos, incluyendo IL-1, FNT-a, IL-6, IL-8, FEC-GM, y factor estimulador de colonias de granulocitos (FEC-G), e inhibir la producción de óxido nítrico. A diferencia de su potente actividad inmunosupresora en los macrófagos, la IL-10 estimula a las células B para proliferar, diferenciarse y producir anticuerpos, y también parece ser un factor de cambio para la producción de IgG4.
En humanos, la IL-10 es un polipéptido de 18 kd codificado por un gen en el cromosoma 1. Muestra importante homología en la secuencia de aminoácidos con una proteína del virus Epstein-Barr que tiene cierta actividad de IL-10 in vitro. No se sabe si la IL-10 viral y de mamífero comparten un mismo gen ancestral. La disfunción inmunológica en el ratón IL-10nulo causa enteritis y colitis. La enfermedad puede transferirse en forma adoptiva a los ratones con IDSC por medio de células T purificadas.39
INTERLEUCINA-12
La IL-12 es un potente estimulador de las células NK, causa la maduración de células T citotóxicas y es un factor de crecimiento para las células T activadas. Por su capacidad para inducir el interferón gama, la IL-12 aumenta la formación de células TH1 e inhibe la formación de células TH2 por la IL-4.40,41
La IL-12 puede proteger a los ratones BALB/c susceptibles contra la infección por L. major cuando reciben antígeno de Leishmania antes o después de la infección. Es posible que pueda emplearse esta citocina como adyuvante en el desarrollo de vacunas.42,42
La estructura de la IL-12 es única entre las citocinas, Esta formada por dos subunidades diferentes producidas por distintos tipos celulares. Una subunidad, IL-12a es una proteína de 35 kd producida por células T y B, células NK y macrofagos, Tiene la estructura de cuatro hélices a semejante a otras citocinas hematopoyéticas. La otra subunidad, IL-12b, es una proteína de 40 kd producida por células B y macrófagos activados. Es homóloga al receptor para IL-6, que contiene la secuencia Trp-Ser-X-Trp-Ser. Por lo tanto, parece ser que el heterodímero de IL-12 está constituido por una proteína semejante a una citocina ligada a una proteína semejante a un receptor de citocina.
INTERLEUCINA-5
La IL-5, producida por células TH2 y células cebadas, estimula el crecimiento y diferenciación de los eosinófilos. Los eosinófilos son activados por la IL-5 y por un factor que aumenta la citotoxicidad de los eosinóficos derivado de monoticos. Los eosinófilos activados destruyen parásitos helmintos y producen daño tisular en estados de alergia. La IL-5, antes denominada factor de crecimiento de células B II y factor de remplazo de células T, estimula el crecimiento y diferenciación de los linfocitos B y, en sinergia con otras citocinas, induce la síntesis de IgM e IgA en células B maduras. La IL-5 es un homodímero de 40 kd relacionado con la familia de las citocinas hematopoyéticas.
INTERLEUCINA-6
La IL-6 es producida por diferentes tipos celulares, incluyendo macrófagos, fibroblastos, células vasculares endoteliales y algunas células T activadas, y tiene un efecto muy amplio sobre diversos tipos celulares.44-46 La IL-6, antes denominada factor estimulador de células B-2 y factor de diferenciación de las células B, induce diferenciación terminal de linfocitos B secretores de inmunoglobulinas, Actúa como un factor de crecimiento para las células plasmáticas malignas. De hecho, se ha observado proliferación policlonal extensa de células plasmáticas en ratones transgénicos que expresan un exceso de Il-6. La producción de IL-6 es estimulada por IL-1 y el FNT-a de los macrófagos, pero la IL-6 inhibe la producción por los macrófagos de IL-1 y FNT-a. La IL-6 actúa sobre las células hepáticas para producir proteínas de fase aguda, como fibrinógeno, alfa2-macroglobulina y proteína de amiloide sérico A.
La IL-6, un péptido que varía en tamaño de 21 a 32 kd por la glucosilación, pertenece a la familia de citocinas hematopoyéticas. En el ratón IL-6nulo existe disrregulación de los progenitores mieloides y eritroides. Como es de esperarse, este ratón no produce proteínas de fase aguda y es más resistente a la infección.
INTERLEUCINA-7
La IL-7, un monómero de glucoproteína de 25 kd, es producida por las células del estroma de la médula ósea y del timo y afecta a las células B tempranas. Induce proliferación de los progenitores B (linfocitos B en los que los genes de las cadenas ligeras y pesadas no están aún rearreglados en la configuración de línea germinal) y de células pre-B (que tienen ya rearreglo del gen de la cadena pesada y contienen cadenas pesadas en el citoplasma) pero no de las células B maduras que producen inmunoglobulina asociada a la membrana (i.e., receptores de células B). En este sentido, la IL-7 debe considerarse como relacionada a otras citocinas hematopoyéticas, como IL-3. La IL-7 estimula también el crecimiento de las células T CD3-, CD4- y CD8- que pueden después responder a IL-2. El timo contiene niveles altos de ARNm para IL-7. La IL-7 puede estimular a las células T maduras y se ha demostrado que aumenta las células T citotóxicas. Se observa un aumento importante en las células pre-B de la médula ósea y linfoices en los ratones transgénicos que expresan IL-7 en exceso.
La IL-7 es un monómero de glucoproteína de 25 kd. Los ratones mutantes deficientes en el receptor de IL-7 tienen una reducción muy importante en la celularidad linfoide tímica y periférica.47
INTERLEUCINA-14
La IL-14, un factor de crecimiento de células B humanas producidas por células malignas B y T, también es conocido como factor de crecimiento de células B de alto peso molecular.48 Induce proliferacion de células B activadas, pero no de células B en reposo e inhibe la secreción de inmunoglobulinas. Los estudios preliminares sugieren que expande en forma selectiva la subpoblaciones de células B de memoria normales.48 Cuando se examinan los leucocitos, solo el linaje de células B tiene receptores para IL-14. La IL-14 es una proteína de 53 kd.
FACTOR INHIBITORIO DE LA MIGRACION
El FIM fue la primera citocina de células T que se descubrió, y se produce después de la estimulación de células T sensibilizadas con el antígeno específico. Esta citocina fue el primer parámetro in vitro de las reacciones de hipersensibilidad tardía [ver figura 6]. Los linfocitos pueden estimularse también en forma inespecífica por mitógenos y complejos inmunes para producir FIM. Se ha clonado el gen para el FIM humano y murino, el monómero es una proteína de 12 kd.49 Además, se ha demostrado que el gen para el FIM es un gen temprano que se expresa en muchos tejidos diferentes.50 Se encuentran grandes cantidades del gen en los macrófagos y las células hipofisiarias. De hecho, el lipopolisacárido (LPS) estimula la liberación de FIM por la hipófisis. Cuando se administra a ratones, el FIM aumenta mucho la letalidad del LPS. Por el contrario, los anticuerpos anti-FIM revierten por completo esta mortalidad.51 Esta citocina participa también en el desarrollo del cristalino ocular.52 Se ha demostrado que el FIM recombinante activa a macrófagos para destruir a Leishmania y los estimula para producir FNT-alfa. Aún no se determina la forma como el FIM interactúa con otras citocinas. El FIM parece ser idéntico al factor inhibitorio de glucosilación, que inhibe la producción de IgE.53

|
| Figura 6 |
| Patrones de migración |
FACTOR DE NECROSIS TUMORAL- a
El FNT-a es una de las sustancias más abundantes producidas por los macrófagos, en especial después de la estimulación por endotoxina. Los macrófagos producen también FNT-a después de la estimulación con interferón gama o FIM. El FNT-a es producido por células T activadas, células NK y células cebadas. Su nombre deriva del hecho de que fue un factor que se encontró en el suero de animales que recibían endotoxina que podía inducir necrosis hemorrágica en los tumores. El FNT-a tiene un papel crucial en la respuesta del huésped a bacterias gram negativas, la fuente de la endotoxina y del LPS. El FNT-a media los efectos del LPS, que en dosis bajas induce una respuesta protectora que causa aumento en la función de los macrófagos, poliactivación de linfocitos B y fiebre. la inmunidad protectora de ciertos organismos intracelulares, como Leishmania, aumenta por efecto del FNT-a. Los efectos antivirales de este factor son causados por la producción de interferón gama. Sin embargo, en concentraciones altas el LPS causa muerte por estado de choque (que puede prevenirse por anticuerpos anti-FNT-a) y coagulación intravascular diseminada. Parte de este fenómeno se conoce como la reacción de Shwartzman. El LPS causa la liberación de FIM, que a su vez aumenta la producción de FNT-a por los macrófagos.
En concentraciones bajas, el FNT-a aumenta la respuesta inflamatoria protectora. Esto induce la expresión de moléculas de adhesión ICAM-1, VCAM-1 y selectina E (ELAM-1) en las células endoteliales vasculares, que promueven la adhesión de neutrófilos, macrófagos y eosinófilos. Estimula también la producción de citocinas por los macrófagos, incluyendo el FNT-a en sí. la IL-1, la IL-6 y diversas quimocinas. El FNT-a aumenta la expresión de moléculas de clase I del CPH y potencia la lisis celular inducida por células T citotóxicas. En concentraciones elevadas el FNT-a puede actuar como una hormona. Funciona como un pirógeno semejante a IL-1 (ver adelante), activa el sistema de coagulación y la producción de proteínas de fase aguda hepáticas y puede causar inmunodeficiencia por la supresión de la médula ósea. El FNT-a inhibe la contractilidad del miocardio al estimular la producción de óxido nítrico y, cuando está presente por tiempo prolongado, causa caquexia. Muchos de los síntomas del paludismo, en especial a nivel cerebral, y algunos de los síntomas de la infección por el virus de inmunodeficiencia humana (VIH) pueden ser mediados por FNT-a.
El FNT-a circula en el suero como homotrímero, formado por subunidades de 17 kd que han surgido de proteolisis de un monómero transmembrana de 25 kd. Se ha evaluado tanto a los anticuerpos anti-FNT-a como a los receptores solubles del FNT-a por su posible papel en el tratamiento de diversos padecimientos inflamatorios.
LINFOTOXINA
Producida en forma exclusiva por las células T activadas, la linfotoxina comparte muchas propiedades biológicas con el FNT-a y también se le conoce como FNT-b. Al igual que el FNT-a, la linfotoxina lisa células tumorales pero no células normales, activa neutrófilos y aumenta la adhesión vascular y la extravasación de los leucocitos. Los efectos de la linfotoxina dependen de la unión con otra proteína, la linfotoxina b, para formar un compleo. Utiliza el mismo receptor celular que el FNT-a.
La linfotoxina es una glucoproteína de 21 a 24 kd. Al contrario del FNT-a, es una proteína secretora que no tiene un componente transmembrana. También se conoce la estructura cristalizada de la linfotoxina unida al dominio extracelular del receptor de FNT de 55 kd.54 El complejo tiene tiene tres moléculas receptoras unidades de modo simétrico a un trímero de linfotoxina. El dominio extracelular del receptor, una estructura muy larga y con ensambles término-terminal de cuatro dominios semejantes entre sí, se une en el surco entre dos subunidades de linfotoxina adyacentes. Esta estructura puede ser representativa de la familia factor de crecimiento neural/receptor de FNT.
INTERLEUCINA-1
La interleucina-1 se produce principalmente por monocitos y macrófagos, pero también por otras células, como células epiteliales y endoteliales. Es un pirógeno endógeno, y muchas de sus funciones son semejantes a las del FNT-alfa. Induce la producción de IL-1 e IL-6 adicional de los macrófagos y también la síntesis de esteroides y liberación de prostaglandina, colagenasa y proteínas de fase aguda. La IL-1 aumenta la expresión de moléculas de superficie en las células endoteliales, causando la adhesión de leucocitos y coagulación, y estimula la producción de quimocinas de macrófagos que a su vez activan neutrófilos. La IL-1 difiere del FNT-alfa en que no produce necrosis de tumores o lesión tisular, aumenta la expresión del complejo principal de histocompatibilidad o, per se, media la reacción de Shwartzman.
La actividad de la IL-1 es resultado de dos polipéptidos, IL-1a e IL-1b, que son producidos por genes separados. La IL-1b es el más abundante. Una proteína de 17 kd, la IL-1b, proviene de la fragmentación de un precursor inactivo de 33 kd por una proteasa de macrófagos. Sin embargo, tanto la molécula IL-1a de 33 kd como la de 17 kd son activas. Los macrófagos producen un antagonista del receptor de IL-1 (IL-1ra) que es estructuralmente semejante a la IL-1 y se une al receptor de IL-1 aunque es inactivo. El IL-1ra, junto con los receptores de IL-1 liberados de las células activadas, inhiben la IL-1 y actúan como reguladores. Estos inhibidores naturales de la IL-1 han sido investigados como agentes potenciales para uso clínico para contrarrestar ciertas proteínas inflamatorias.
INTERLEUCINA-8 Y OTRAS QUIMOCINAS
La IL-8 y otras citocinas quimiotácticas o quimocinas, son importantes para inducir y mantener las reacciones inflamatorias. Diferentes quimocinas pueden unirse al mismo receptor. El peso molecular de las quimocinas varía de 8 a 11 kd, y están clasificadas en dos subfamilias, a y b.
Las moléculas de la subfamilia de quimocinas a se distinguen por su localización en el cromosoma 4 y por su secuencia de aminoácidos, en la que las dos primeras cisteínas están separadas por un aminoácido (Cys-X-Cys). La subfamilia a incluye IL-8, b-tromboglobulina (b-TG), gro-a, gro-b, gro-g y factor plaquetario 4. Suelen ser producidos por monocitos, algunos son producidos por otras células, incluyendo células T, células endoteliales y plaquetas. La IL-8 induce la formación de integrinas de unión a neutrófilos en las células endoteliales, causando acúmulo rápido de neutrófilos en los tejidos. La quimocina gro, también conocida como MIP-2a y MIP-2b, causa también acúmulo de neutrófilos, así como liberacipde enzimas lisosomales que contribuyen a la respuesta inflamatoria local. El factor plaquetario 4 y la b-TG, liberadas de plaquetas agregadas, estimulan a los fibroblastos requeridos para la reparación en sitios de hemorragia o trombosis.
Las moléculas de la subfamilia de quimocinas b se localizan en el cromosoma 17 y tienen un patrón aminoácido cisteína-cisteína (Cys-Cys). Incluyen a RANTES (quimocina expresada y secretada en las células T normales y que regula su activación), al factor quimiotáctico y activador de macrófagos (FQAM), a MIP-2a y a MIP-2b. Las quimocinas de la subfamilia b son producidas en monocitos y células T activadas. RANTES es un atractor potente para células T de memoria, pero no para células T nativas, y también atrae monocitos. El FQAM actúa solo en los monocitos, atrayendo, activándolos, y regulando la expresión de integrinas en su superficie. MIP-1a y MIP-1b atraen solo monocitos.
LINFOTACTINA
La linfotactina, un factor quimiotáctico de linfocitos, fue clonado recientemente de timocitos tempranos (i.e., células pro-T). Este factor puede atraer a los progenitores celulares al timo.55
FACTOR ESTIMULADOR DE COLONIAS DE GRANULOCITOS Y MACROFAGOS
El FEC-GM actúa sobre células de la médula ósea que ya están comprometidas para convertirse en leucocitos. Es producido por células T, macrófagos, fibroblastos y células endoteliales vasculares. Los macrófagnos de la médula ósea cultivados con FEC-GM tienen mayor capacidad para presentar antígenos a células TH1, comparado con los macrófagos de la médula ósea cultivados en factor estimulador de colonias de macrófagos (FEC-M).56 La inyección de macrófagos de médula ósea cultivados con FEC-GM con antígeno de L. major estimula la respuesta de células TH1 en los ratones susceptibles y los hace resistentes al reto con L. major. También se ha encontrado que que el FEC-GM activa macrófagos.57 El FEC-GM es un dímero de 22 kd que no circula sino que actúa en forma local. Se ha usado para contrarrestar la leucopenia en pacientes tratados con quimioterapia.
El FEC-GM es un miembro de la familia de las citocinas hematopoyéticas. Otras citocinas que son principalmente hematopoyéticas incluyen al factor de células tronco, la IL-3, el FEC-M, el FEC-G, la IL-9 y la IL-11.
Papel de los carbohidratos en la regulación de las respuestas de las células T
Estudios recientes sobre la respuesta inmunológica a los parásitos Schistosoma han demostrado que un carbohidrato de superficie, lacto-N-fucopentaosa-III (LNFP-III), puede cambiar una respuesta mediada por células TH1 en una respuesta TH2 mediada principalmente por anticuerpos.58 El LNFP-III estimula las células B convencionales y las células B1 para secretar grandes cantidades de IL-10. La IL-10, al actuar a través de los macrófagos, inhibe las células TH1 y favorece la respuesta TH2.
Esto tiene implicaciones importantes para la imunidad a muchos organismos porque este azúcar, que contiene Lewisx, está presente en otros parásitos y células neoplásicas. El microrganismo o célula neoplásica puede usar el carbohidrato para inhibir la respuesta inmunológica mediada por células protectora del huésped. Este dato puede permitir el desarrollo de una estrategia novedosa para la prevención de la evasión inmunológica por microrganismos y células tumorales, por ejemplo, bloqueando el receptor ß1 usado por el LNFP-III, o el desarrollo de respuestas de anticuerpos más intensas usando LNFP-III en la vacuna. Aún más, recientemente se ha demostrado que la misma familia de oligosacáridos se une a la proteína NKR-P1 de las células NK. Esta interacción es crucial para el reconocimiento de las células blanco por las células NK y para la citotoxicidad.59
Activación de las células B
Cuando un receptor de la célula B es activado por un antígeno soluble, ocurre uno de dos eventos: apoptosis o proliferación y maduración posterior de las células B.60 Las señales para estos procesos se generan dentro de las células por la Iga e Igb, dos proteínas que se asocian con los receptores de la célula B en la misma forma que las proteínas CD3 se asocian con el RCT. La Iga y la Igb reclutan moléculas de transducción de señales en una forma semejante a la del complejo RCT-CD3, específicamente en dos fases de cinasas de tirosina, la primera consistente en p56lyn, p59fyn y p55blk, y la segunda en p72syk. Se reclutan entonces algunas de las mismas proteínas descritas para la activación de la célula T [ver antes, Transducción de señales en Reconocimiento de antígenos por las células T], por ejemplo PI3-cinasa, p95vav, p21ras, p72-74raf, cinasas mek, PLC-b y PLC-g). La vía de activación de las células B es semejante pero no idéntica a la de activación de las células T.
Se sabe relativamente poco sobre la señalización que tiene lugar en las células B cuando presentan un antígeno a una célula T que lo reconoce. Como cuando se activan por antígenos solubles, la interacción célula T-B causa apoptosis o proliferación de las células B. Sin embargo, con la ayuda de la célula T la proliferación no provoca la aparición de diferentes clases de células B.61
CELULAS FOLICULARES EN MANTO
Las células foliculares en manto expresan IgD (IgD+, CD38-) en su superficie y pueden madurar a células plasmáticas que producen solo anticuerpos de IgM. Estas células producen IgM que muestra gran heterogeneidad, pero las células no sufren mutación somática para generar un número aún mayor de moléculas de anticuerpos heterogéneos.
CENTROCITOS
Los procesos que causan mutación somática ocurren en los centrocitos, que son células B IgD-, CD38+ que han sufrido cambio de clase para producir IgG. El paso de maduración de las células B IgD+, CD38- a células B IgD- CD38+ depénde del contacto intercelular con células T antígeno específicas [ver figura 7] y con citocinas. La IL-4 tiene especial importancia para los eventos que causan el cambio de clase.
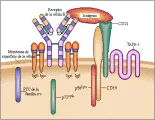
|
| Figura 7 |
| Correceptores en las células B |
CELULAS DE MEMORIA
La célula B más madura es la célula de memoria, que no expresa IgD o CD38, pero que se distingue por otros marcadores de superficie y por su localización dentro de los centros germinales. Las células de memoria pueden convertirse en células plasmáticas que producen IgM, IgG, IgA e IgE. Sin embargo, en general la generación de células plasmáticas es independiente de la ayuda de los linfocitos T.
INTERACCION DEL LIGANDO CD40-CD40
Recientemente se ha aclarado la importancia de la interacción entre el ligando CD40 y CD40 (CD40L).62 El CD40L es una glucoproteína tipo 2 que se localiza en la superficie de la célula T relacionada al FNT. La expresión temporal de CD40L en la superficie de las células T es inducida por la unión del RCT con el complejo antígeno-molécula del CPH y por la unión de moléculas B7 con CD28 o CTLA-4. A su vez, la ligadura de CD40 por CD40L causa estimulación del cambio de clase de inmunoglobulinas en las células B. Esto se demostró mejor por el descubrimiento del defecto molecular en una inmunodeficiencia genética denominada síndrome de hiper-IgM. La mutación del gen CD40L es responsable del cambio de clase defectuoso que sucede en este síndrome. Las respuestas de las células B independientes de células T y las respuestas inducidas por el anti-CD40 no se afectan por esta mutación. Por lo tanto, la ayuda de la célula T para activar al linfocito B se realiza principalmente por la vía coestimuladora CD40-CD40L. Los pacientes con síndrome de hiper-IgM también son susceptibles a los patógenos que normalmente son atacados por las células T, produciendo infecciones oportunistas semejantes a las del SIDA, lo que destaca el papel de la coestimulación CD40L en la activación de las células T normales. La ausencia de CD40L tiene un efecto más dramático en el cambio de clase de Ig que la ausencia de células T. Por ello, la suposición lógica es que CD40L podría expresarse en otras células que no son T, pudiendo inducir el cambio de clase en las células B a través de la unión con CD40.
Inmunidad mediada por células
La inmunidad mediada por células consiste en la destrucción de microrganismos invasores como bacterias, virus, hongos y parásitos, la destrucción de células tumorales, el rechazo de injertos tisulares y la lesión a los tejidos en varios estados patológicos, incluyendo la autoinmunidad. Las reacciones inmunes mediadas por células pueden también inducirse por contacto con antígenos, como los encontrados en la hiedra venenosa y muchos medicamentos. Los medicamentos provocan más reacciones mediadas por células cuando se aplican por vía tópica más que sistémica.
La mayoría de las reacciones inmunes mediadas por células incluyen la interacción inicial entre células T sensibilizadas y antigenos en las células presentadoras. Esta reacción puede desencadenar varias vías efectoras, incluyendo activación de células T citotóxicas, estimulación de la producción de citocinas que activan macrófagos y promueven la proliferación de células NK, y producción de anticuerpos incluidos en la CMCDA por células NK y otras células. Aunque las reacciones inmunes mediadas por células diferentes a la CMCDA no requieren la presencia de anticuerpos o complemento, pueden modificarse por estos factores humorales. Los eventos subsecuentes requieren de la cooperación entre los diferentes subtipos de células T. Las reacciones incluidas son controladas por diversas citocinas.
Los mecanismos de inmunidad mediada por células incluyen interacciones entre las células T y macrófagos que pueden ser protectoras (causando destrucción del microrganismo invasor) y dañinas (provocando inflamación y destrucción tisular). En ocasiones, ambas respuestas van de la mano, por ejemplo, en la tuberculosis, tanto la destrucción del bacilo tuberculoso como la formación de cavernas pulmonares son consecuencia de la interacción célula T-macrófago.
CELULAS T CITOTOXICAS
Las células T citotóxicas son células efectoras antígeno-específicas que son importantes para resistir a los agentes infecciosos, en especial a virus existentes en células que no son macrófagos, destrucir tumores y en el rechazo de injertos. La mayoría de las células T citotóxicas con CD8+, y reconocen antígenos presentados por moléculas de clase I del CPH, aunque algunas pocas son T CD4+ y reconocen al antígeno en presencia de moléculas de clase II del CPH. Las células T citotóxicas se desarrollan y sufren selección en el timo, después maduran a células T citotóxicas funcionales en los tejidos. El precursor inmaduro de las células T citotóxicas tiene RCT de superficie y expresa moléculas CD8 pero no es funcional. La células precursora adquiere capacidad lítica después de dos señales. La primera consiste en activación por contacto con el complejo antígeno-molécula del CPH en una CPA. La segunda señal es proporcionada por citocinas, generalmente producida por células T cooperadoras CD4+. Entre las muchas citocinas que se piensa participan en la activación de las células T citotóxicas se incluyen el interferón gama, la IL-2, la Il-4, la Il-7 y la IL-12. Los detalles de la activación de las células T citotóxicas y las moléculas de adhesión que participan se describen en otra parte [ver antes, Procesamiento y presentación del antígeno a las células T]. Las células T citotóxicas desarrollan gránulos que contienen moléculas citotóxicas, incluyendo perforinas (proteínas que producen orificios o poros en la membrana de superficie de la célula), granzima A, granzima B y esterasas de serina. De éstas, las perforinas son las más importantes, como se ha demostrado en ratones en el que se delecionó el gen de la perforina.63 Un segundo mecanismo de destrucción celular incluye la unión del ligando Fas en la célula T citotóxica y la molécula Fas en la célula blanco.64 Este es el único mecanismo disponible para el ratón perforinanulo, y se usa en forma preferencial (aunque no exclusiva) en las células T citotóxicas CD4+. Las células T citotóxicas también producen citocinas, como linfotoxina, interferón gama y FNT-a.
La capacidad de las células T citotóxicas para destruir células blanco infectadas por virus está gobernada por restricciones antigénicas y genéticas. Los linfocitos de ratones que han sido infectados por un virus matarán solo a las células blanco que hayan sido infectados con el mismo virus y porten el mismo antígeno de clase I del CPH. Por lo tanto, para que ocurra la lisis, las células blanco deben portar tanto el mismo antígeno viral como la misma molécula de clase I del CPH que las células que indujeron en forma original a la células T citotóxicas. Los antígenos de clase I del CPH parecen ser los ideales para activar a las células T citotóxicas porque están presentes en todas las células excepto en los eritrocitos. Los estudios han demostrado que las células T citotóxicas pueden ser la principal vía efectora de la inmunidad mediada por células contra ciertos virus en vivo, más que los macrófagos activados por citocinas. Los estudios in vitro han demostrado que si las células blanco se infectan con un virus, pueden ser destruidas después de algunas horas por las células T citotóxicas específicas, antes de que el virus tenga oportunidad de replicarse. Un subgrupo de células T citotóxicas que tienen RCT-gd puede destruir sin restricción del CPH.
CELULAS ASESINAS NATURALES
Las células NK son linfocitos granulares grandes que portan diversos marcadores de superficie celular, incluyendo NK1.1 (un receptor que se pensaba participaba en el reconocimiento de la célula blanco), CD2 y CD16 (un receptor de Fc IgG de baja afinidad, FcgRIII). Las células NK no tienen el marcador complejo RCT-CD3 ni la inmunoglobulina de superficie de las células B. Estas células pueden destruir células infectadas por virus y diversas líneas celulares tumorales in vitro, y se piensa que tienen actividades antivirales y antitumorales. Predominan en la respuesta a la reacción injerto contra huésped después del trasplante de médula ósea. Las células NK no requieren sensibilización previa para volverse citotóxicas, y sus actividades no están restrigidas por el CPH. CD3-s, que funciona en la transducción de señales, se asocia con CD16 en las células NK, y puede detectarse CD3-e en las células NK activadas. En general, se piensa que las células T y las células NK pueden tener un precursor común.
Las células NK no maduran solo en el timo. De hecho, pueden activarse en el ratón deficiente de timo y en el ratón con DICS. La IL-12 es un inductor potente de la proliferación y activación de las células NK, también el interferón gama y la IL-2 activan estas células. La IL-2 aumenta la capacidad tumoricida de las células NK que causa el desarrollo de células LAK. Las células NK pueden también secretar interferón gama, que actúa en forma autócrina para incrementar la actividad de las células NK. El FNT parece contrarrestar el desarrollo de las células NK inducido por IL-12.
Las células NK se fijan a su célula blanco por un receptor en un paso independiente de Ca2+ [ver figura 8]. Sin embargo, una vez que las células NK se activan, requieren iones Ca2+ para la lisis. Esto se logra por orientación de sus gránulos hacia la célula blanco y la liberación de perforinas y enzimas proteolíticas. Destruyen por mecanismos semejantes a los de las células T citotóxicas, como se ha demostrado en el ratón perforinanulo. La destrucción por células NK, al contrario de la destrucción por células T citotóxicas, no es bloqueada por anticuerpos monoclonales anti-CD3 o anti-CD8, pero sí por anticuerpos contra LFA-1.
|
|
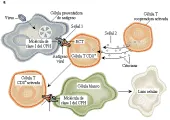
Las células NK, macrófagos, neutrófilos y eosinófilos se encuentran entre las células que median las reacciones de CMCDA. El anticuerpo, por lo general IgG, se une al receptor Fc en la célula efectora y el sitio de combinación del anticuerpo se une al antígeno en la célula blanco, lo que produce una reacción antígeno específica [ver figura 8]. Por ejemplo, los eosinófilos son eficaces para destruir esquistosómulas cubiertas por anticuerpos, la larva del parásito de Schistosoma. El mecanismo de destrucción incluye la unión de los eosinófilos a la larva a través del anticuerpo unido, seguido de la liberación de proteínas básicas y otras moléculas de los gránulos eosinófilos, causando la formación de poros en la célula blanco. Además de la IgG, la IgE y la IgA pueden mediar también la CMCDA. Los eosinófilos son reclutados a través de la producción de IL-5 por las células T. Los eosinófilos muestran mayor actividad citotóxica después de su activación por citocinas, como el factor estimulador de eosinófilos citotóxicos (producido por macrófagos), el FNT-a, el FEC-GM y la IL-5.
MACROFAGOS ACTIVADOS
Los macrófagos activados son células efectoras importantes en las respuestas inmunes mediadas por células contra varios microrganismos, incluyendo M. tuberculosis, M. leprae, Listeria monocytogenes, Legionella pneumophila, Salmonella, Leishmania y Toxoplasma gondii.65 En esta vía efectora, el macrófago ingiere al microrganismo y después presenta el péptido antigénico procesado en el surco de la molécula de clase II del CPH a la célula T cooperadora CD4+ [ver figura 8]. En la presentación subsecuente al mismo antígeno por el mismo macrófago, las células T cooperadoras sensibilizadas son activadas para producir y secretar varios factores quimiotácticos que atraen más monocitos/macrófagos, granulocitos y linfocitos al sitio de la reacción. Las propiedades de los macrófagos se alteran por FAM, muchos de los cuales son producidos por células T, incluyendo interferón gama (la citocina FAM más importante, así como FEC-GM, FIM, IL-3, IL-4 y FNT-a. El LPS, al actuar con algunas de estas citocinas, aumenta también la activación de los macrófagos [ver figura 9]. La activación de los macrófagos puede ser modulada y disminuida por otras citocinas, incluyendo FTC-b, IL-4 e IL-10, y por prostaglandinas.66
|
| Figura 9 |
| Acción del lipopolisacárido bacteriano |
Los FAM inducen la expresión de muchos genes en los macrófagos, causando el fenotipo de activación. Aumenta la fagocitosis y la endocitosis. La mayor expresión de HLA-DR, ICAM-1 y LFA-3 y la inducción de B7 aumenta la presentación del antígeno. La mayor afinidad de los receptores de Fc IgG permite una mejor opsonización de los microrganismos. Los macrófagos activados destruyen microrganismos y células tumorales a través de diversos mediadores, incluyendo la mayor producción de sintetasa de óxido nítrico, lo que causa generación de óxido nítrico y producción de peróxido de hidrógeno y otros radicales reactivos del oxígeno.
La capacidad de los macrófagos activados para destruir microrganismos aumenta en forma inespecífica, esto significa que los macrófagos pueden eliminar no solo al organismo específico que indujo la producción del FAM, sino tambien a otros organismos. Por ejemplo, un macrófago que ha sido activado para destruir L. monocytogenes puede también destruir organismos Salmonella de modo eficaz.
Los macrófagos activados producen muchos mediadores inflamatorios que pueden aumentar la inmunidad atrayendo leucocitos al área, pero también pueden promover la inflamación que causa destrucción tisular. Las sustancias producidas por los macrófagos activados incluyen prostaglandinas, quimocinas, factor activador de plaquetas, factores de coagulación, factor activador del plasminógeno, factor de crecimiento derivado de plaquetas y factor de crecimiento de fibroblastos. Estos factores participan en la inflamación, la coagulación, la proliferación de fibroblastos, la producción de colágena, la fibrosis y la angiogénesis. Las consecuencias patológicas de esta interacción célula T-macrófago es la base de las reacciones de hipersensibilidad tardía.
REACCIONES DE HIPERSENSIBILIDAD TARDIA
Las reacciones de hipersensibilidad tardía están mediadas por células T. Estas reacciones alcanzan su máximo 24 a 48 horas después de la introducción de un antígeno en la piel, a diferencia de la hipersensibilidad alérgica inmediata mediada por IgE, que ocurre en segundos a minutos después de la introducción de un antígeno, y de las reacciones mediadas por complejos inmunes-IgG-complemento (de Arthus), que alcanzan su máximo después de dos a cuatro horas. A diferencia de las reacciones inmediatas y de Arthus que pueden transferirse a los animales humanos por anticuerpos, las reacciones de hipersensibilidad tardía se transfieren solo por células T sensibilizadas.
El prototipo de la reacción de hipersensibilidad tardía es la prueba cutánea a la tuberculina. Cuando se inyecta derivado proteico purificado (PPD) de M. tuberculosis en la piel de una persona que se ha sensibilizado antes por una infección previa, el antígeno es procesado por CPA, incluyendo células endoteliales venulares y se presenta a las células T sensibilizadas con PPD. Esto permite la elaboración de citocinas, la activación de algunos macrófagos y la producción y liberación de más mediadores, como quimocinas, FNT-? e interferón gama. Estos mediadores atraen leucocitos e inducen la expresión secuencial de las moléculas de adhesión vascular ELAM-1, VCAM-1 e ICAM-1, provocando reclutamiento y extravasación de leucocitos en el área [ver figura 10]. Aunque los neutrófilos aparecen pronto en las reacciones de hipersensibilidad tardía, después de 24 horas la mayoría de las células son macrófagos, con algunos linfocitos (la radiación en dosis bajas, que destruye principalmente a los precursores de macrófagos en la médula ósea, disminuye en forma importante el infiltrado celular en la hipersensibilidad tardía) [ver figura 11]. Las lesiones por hipersensibilidad tardía característicamente son firmes o induradas como resultado del depósito de fibrina en la lesión. La hipersensibilidad por contacto, como la observada con la hiedra venenosa o las alergias a medicamentos, es una forma de hipersensibilidad tardía. En algunos casos las lesiones aparecen antes que en la hipersensibilidad tardía del tipo tuberculina, y contienen muchos basófilos. A esta reacción se denomina hipersensibilidad basófila cutánea o de Jones-Mote. Los mecanismos que participan en las reacciones de hipersensibilidad tardía también se observan en procesos granulomatosos y en algunas reacciones autoinmunes.
|
|
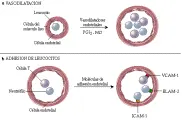
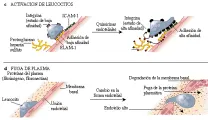
|
| Figura 11 |
| Histología de la reacción de hipersensibilidad |
Bibliografía
-
Nossal GJV: Current concepts: immunology: the basic components
of the immune system. N Engl J Med 316:1320, 1987
-
Engelhard VH: How cells process antigens. Sci Am 271(2):54,
1994
-
Stern LJ, Brown JH, Jardetzky TS, et al: Crystal structure
of the human class II MHC protein HLA-DR1 complexed with an influenza virus
peptide. Nature 368:215, 1994 [PMID
8145819]
-
Brown JH, Jardetzky TS, Gorga JC, et al: Three-dimensional
structure of the human class II histocompatibility antigen HLA-DR1. Nature
364:33, 1993 [PMID
8316295]
-
Jardetzky TS, Brown JH, Gorga JC, et al: Three-dimensional
structure of a human class II histocompatibility molecule complexed with
superantigen. Nature 368:711, 1994 [PMID
8152483]
-
Rock KL, Gramm C, Rothstein L, et al: Inhibitors of the proteasome
block the degradation of most cell proteins and the generation of peptides
presented on MHC class I molecules. Cell 78:761, 1994 [PMID
8087844]
-
Amigorena S, Drake JR, Webster P, et al: Transient accumulation
of new class II MHC molecules in a novel endocytic compartment in B lymphocytes.
Nature 369:113, 1994 [PMID
8177316]
-
Tulp A, Verwoerd D, Dobberstein B, et al: Isolation and characterization
of the intracellular MHC class II compartment. Nature 369:120, 1994 [PMID
8177317]
-
Morris P, Shaman J, Attaya M, et al: An essential role for
HLA-DM in antigen presentation by class II major histocompatibility molecules.
Nature 368:551, 1994 [PMID
8139689]
-
Fling SP, Arp B, Pious D: HLA-DMA and -DMB genes are both
required for MHC class II/peptide complex formation in antigen-presenting
cells. Nature 368:554, 1994 [PMID
8139690]
-
Sanderson F, Kleijmeer MJ, Kelly A, et al: Accumulation of
HLA-DM, a regulator of antigen presentation, in MHC class II compartments.
Science 266:1566, 1994 [PMID
7985027]
-
Beckman EM, Porcelli SA, Morita CT, et al: Recognition of
a lipid antigen by CD1-restricted ab+
T cells. Nature 372:691, 1994 [PMID
7527500]
-
Spits H, van Schooten W, Keizer H, et al: Alloantigen recognition
is preceded by nonspecific adhesion of cytotoxic T cells and target cells.
Science 232:403, 1986 [PMID
3485822]
-
Matsui K, Boniface JJ, Reay PA, et al: Low affinity interaction
of peptide-MHC complexes with T cell receptors. Science 254:1788, 1992
-
Springer T: Traffic signals for lymphocyte recirculation
and leukocyte emigration: the multistep paradigm. Cell 76:301, 1994
-
Clark EA, Ledbetter JA: How B and T cells talk to each other.
Nature 367:425, 1994 [PMID
8107800]
-
Weiss A, Littman DR: Signal transduction by lymphocyte antigen
receptors. Cell 76:263, 1994
-
Terhorst CP, Staal F, Spits H, et al: T lymphocyte signal
transduction. Molecular Immunology (Frontiers in Molecular Biology series),
2nd ed. Hames BD, Glover DM, Eds. IRL Press, Washington, DC, 1995
-
Arpaia E, Shahar M, Dadi H, et al: Defective T cell receptor
signaling and CD8+ thymic selection in humans lacking Zap-70
kinase. Cell 76:947, 1994 [PMID
8124727]
-
Elder ME, Lin D, Clever J, et al: Human severe combined immunodeficiency
due to a defect in ZAP-70, a T cell tyrosine kinase. Science 264:1596,
1994 [PMID
8202712]
-
Hutchcroft JE, Bierer BE: Activation-dependent phosphorylation
of the T-lymphocyte surface receptor CD28 and associated proteins. Proc
Natl Acad Sci USA 91: 3260, 1994 [PMID
7512728]
-
Chan IT, Limmer A, Louie MC, et al: Thymic selection of cytotoxic
T cells independent of CD8a-Lck association.
Science 261:1581, 1993 [PMID
8372352]
-
Paul WE, Seder RA: Lymphocyte responses and cytokines. Cell
76:241, 1994 [PMID
7904900]
-
De Vos AM, Ultsch M, Kossiakoff AA: Human growth hormone
and extracellular domain of its receptor: crystal structure of the complex.
Science 255:306, 1992 [PMID
1549776]
-
Kondo M, Takeshita T, Ishii N, et al: Sharing of the interleukin-2
(IL-2) receptor g chain between receptors for
IL-2 and IL-4. Science 262:1874, 1993 [PMID
8266076]
-
Russell SM, Keegan AD, Harada N, et al: Interleukin-2 receptor
g chain: a functional component of the interleukin-4
receptor. Science 262:1880, 1993 [PMID
8266078]
-
Wlodawer A, Pavlovsky A, Gustchina A: Hematopoietic cytokines:
similarities and differences in the structures, with implications for receptor
binding. Protein Science 2: 1373, 1993 [PMID
8401223]
-
Kundig TM, Schorle H, Bachmann MF, et al: Immune responses
in interleukin-2-deficient mice. Science 262:1059, 1993 [PMID
8235625]
-
Simpson SJ, Mizoguchi E, Bhan A, et al: Evidence that CD4+
but not CD8+ T cells cause colitis in IL-2-deficient mice. Eur
J Immunol (in press)
-
Grabstein KH, Eisenman J, Shanebeck K, et al: Cloning
of a T cell growth factor that interacts with the b
chain of the interleukin-2 receptor. Science 264:965, 1994
-
Kuhn R, Rajewsky K, Muller W: Generation and analysis of
interleukin-4 deficient mice. Science 254:707, 1991 [PMID
1948049]
-
Kopf M, Le Gros G, Bachmann M, et al: Disruption of the murine
IL-4 gene blocks Th2 cytokine responses. Nature 362:245, 1993 [PMID
8384701]
-
Zurawski G, de Vries JE: Interleukin 13, an interleukin 4-like
cytokine that acts on monocytes and B cells, but not on T cells. Immunol
Today 15(1):19, 1994
-
Badaro R, Falcoff E, Badaro FS, et al: Treatment of visceral
leishmaniasis with pentavalent antimony and interferon gamma. N Engl J
Med 322:16, 1990 [PMID
2104665]
-
Shull MM, Ormsby I, Kier AB, et al: Targeted disruption of
the mouse transforming growth factor-b1 gene
results in multifocal inflammatory disease. Nature 359:693, 1992 [PMID
1436033]
-
Moore KW, O'Garra A, de Waal Malefyr R, et al: Interleukin-10.
Ann Rev Immunol 11:165, 1993
-
Yssel H, Johnson KE, Schneider PV, et al: T cell activation-inducing
epitopes of the house dust mite allergen Der p I. J Immunol 148:738,
1992 [PMID
1370514]
-
Parronchi P, Macchia D, Piccinni M-P, et al: Allergen- and
bacterial antigen-specific T-cell clones established from atopic donors
show a different profile of cytokine production. Proc Natl Acad Sci USA
88:4538, 1991 [PMID
1827920]
-
Kuhn R, Lohler J, Rennick D, et al: Interleukin-10-deficient
mice develop chronic enterocolitis. Cell 75:263, 1993 [PMID
8402911]
-
Wolf SF, Temple PA, Kobayashi M, et al: Cloning of cDNA for
natural killer cell stimulatory factor, a heterodimeric cytokine with multiple
biologic effects on T and natural killer cells. J Immunol 146:3074, 1994
-
Brunda MJ: Interleukin-12. J Leukoc Biol 55:280, 1994
-
Alfonso LCC, Scharton TM, Vieira LQ, et al: The adjuvant
effect of interleukin-12 in a vaccine against Leishmania major.
Science 263:235, 1994
-
Heinzel FP, Schoenhaut DS, Rerko RM, et al: Recombinant interleukin
12 cures mice infected with Leishmania major. J Exp Med 177:1505,
1993 [PMID
8097524]
-
Bernad A, Kopf M, Kulbacki R, et al: Interleukin-6 is required
in vivo for the regulation of stem cells and committed progenitors of the
hematopoietic system. Immunity 1:725, 1994
-
Kopf M, Lamers M, Bluethman H, et al: IL-6 deficient mice
show immunologic abnormalities. Nature 376:339, 1994
-
Poli V, Balena R, Fattori E, et al: Interleukin-6 deficient
mice are protected from bone loss caused by estrogen depletion. EMBO J
13:1189, 1994 [PMID
8131749]
-
Peschon JJ, Morrissey PJ, Grabstein KH, et al: Early lymphocyte
expansion is severely impaired in interleukin 7 receptor-deficient mice.
J Exp Med 180:1955, 1994 [PMID
7964471]
-
Ambrus JL Jr, Pippin J, Joseph A, et al: Identification
of a cDNA for a human high-molecular-weight B-cell growth factor. Proc
Natl Acad Sci USA 90:6330, 1993 [PMID
8327514]
-
Weiser WY, Temple PA, Witek-Giannotti JS, et al: Molecular
cloning of a cDNA encoding a human macrophage migration inhibitory factor.
Proc Natl Acad Sci USA 86:7522, 1989 PMID
2552447]
-
Lanahan A, Williams JB, Sanders LK, et al: Growth factor-induced
delayed early response genes. Mol Cell Biol 12:3919, 1992 [PMID
1508193]
-
Bernhagen J, Calandra T, Mitchell RA, et al: MIF is a pituitary-derived
cytokine that potentiates lethal endotoxaemia. Nature 365:756, 1993 [PMID
8413654]
-
Wistow GJ, Shaughnessy MP, Lee DC, et al: A macrophage migration
inhibitory factor is expressed in the differentiating cells of the eye
lens. Proc Natl Acad ci USA 90:1272, 1993
-
Lui Y-C, Nakano T, Elly C, et al: Requirement of posttranslational
modifications for the generation of biologic activity of glycosylation-inhibiting
factor. Proc Natl Acad Sci USA 91:11227, 1994
-
Banner DW, D'Arcy A, Janes W, et al: Crystal structure of
the soluble human 55 kd TNF receptor-human TNFb
complex: implications for TNF receptor activation. Cell 73:431, 1993 [PMID
8387891]
-
Kelner GS, Kennedy J, Bacon KB, et al: Lymphotactin: a cytokine
that represents a new class of chemokine. Science 266:1395, 1994 [PMID
7973732]
-
Germann T, Mattner F, Partenheimer A, et al: Different accessory
function for T H1 cells of bone marrow derived macrophages cultured
in granulocyte macrophage colony stimulating factor or macrophage colony
stimulating factor. Int Immunol 4:755, 1992 [PMID
1386751]
-
Doherty TM, Coffman RL: Leishmania antigens presented
by GM-CSF-derived macrophages protect susceptible mice against challenge
with Leishmania major. J Immunol 150:5476, 1993
-
Velupillai P, Harn DA: Oligosaccharide-specific induction
of interleukin 10 cells from schistosome-infected mice: a mechanism for
T-cell subsets. Proc Natl Acad Sci USA 91:18, 1994 [PMID
7904066]
-
Bezouska K, Yuen C-T, O'Brien J, et al: Oligosaccharide ligands
for NKR-P1 protein activate NK cells and cytotoxicity. Nature 372:150,
1994 [PMID
7969447]
-
B-cell signaling special. Immunol Today 15:393, 1994
-
Pascual V, Liu Y-J, Magalski A, et al: Analysis of somatic
mutation in five B cell subsets of human tonsil. J Exp Med 180:329, 1994
[PMID
8006591]
-
Durie FH, Foy TM, Masters SR, et al: The role of CD40 in
the regulation of humoral and cell-mediated immunity. Immunol Today 15:406,
1994 [PMID
7524518]
-
KŠgi D, Ledermann B, Bürki K, et al: Cytotoxicity
mediated by T cells and natural killer cells is greatly impaired in perforin-deficient
mice. Nature 369:31, 1994 [PMID
8164737]
-
KŠgi D, Vignaux F, Ledermann B, et al: Fas and perforin pathways
as major mechanisms of T cell-mediated cytotoxicity. Science 265:528, 1994
[PMID
7518614]
-
Paul WE: Infectious diseases and the immune system. Sci Am
269(3):57, 1993
-
Bogdan C, Nathan C: Modulation of macrophage function by
transforming growth factor b, interleukin-4,
and interleukin-10a. Ann N Y Acad
Sci 685:713, 1993 [PMID
8363277]