Inmunología
⭳ Abrir artículo (PDF)662.8 KBEste artículo fue revisado respecto a la Edición 3/2000. Ver esa versión →
Contenido del artículo
I ORGANOS Y CELULAS DEL SISTEMA INMUNE
- Características del sistema inmune
- Organos linfoides y circulación de linfocitos
- ORGANOS LINFOIDES PRIMARIOS
- ORGANOS LINFOIDES SECUNDARIOS
- OTROS TEJIDOS LINFOIDES
- CIRCULACION DE LINFOCITOS
- Células inmunocompetentes de la respuesta inmunológica
I ORGANOS Y CELULAS DEL SISTEMA INMUNE
DR. JOHN DAVID
Características del sistema inmune
Por la capacidad de distinguir lo propio de lo que no lo es, el sistema inmune es el mediador de la relación del individuo con el medio ambiente que le rodea. Cuando el sistema inmune funciona de manera apropiada, protege al organismo de las infecciones; cuando no, los fallos del sistema inmune producen algunas de las enfermedades más graves y difíciles que el médico puede encontrar en la práctica médica.
MEMORIA Y ESPECIFICIDAD INMUNOLOGICA
La experiencia con las infecciones ha demostrado que el organismo tiene memoria para enfermedades tales como la viruela, el sarampión o la parotiditis. Una vez que un individuo normal ha padecido una enfermedad como las mencionadas, el sistema inmune la reconoce y previene su recurrencia. Además de memoria, el sistema tiene especificidad, esto es, tiene la extraordinaria capacidad para discriminar entre diferentes antígenos, aun cuando sus estructuras estén estrechamente relacionadas entre sí.
Existen dos grupos principales de linfocitos, los linfocitos T (también llamado linfocitos dependientes del timo o células T) y los linfocitos B (también conocidos como linfocitos derivados de la médula ósea o células B). La capacidad de los linfocitos T y de los anticuerpos producidos por los linfocitos B para discriminar entre los antígenos está determinada por los genes de la región variable (V). Durante la maduración de los linfocitos T y B ocurre rearreglo de las secuencias de ADN en estos genes de la región V, además de mutaciones somáticas adicionales en las células B. Los varios cientos de genes de la región V que se forman codifican las inmunoglobulinas (anticuerpos) y los receptores de las células T (RCT), que pueden distinguir alrededor de un billón de diferentes especificidades antigénicas.
Existe un número enorme de linfocitos, cada uno de los cuales tiene un receptor de superficie que reconoce un solo determinante antigénico o epítope. Después de interactuar con un antígeno, los linfocitos T proliferan y se diferencían en células de memoria y en células que regulan la producción de anticuerpos por los linfocitos B y las respuestas efectoras mediadas por células de otros linfocitos T. Después de interactuar con un antígeno, los linfocitos B proliferan y se diferencían en células de memoria y células plasmáticas, encargadas de la producción de anticuerpos.
Antes de ser reconocido por las células T, el antígeno es captado por una célula presentadora de antígeno (CPA) (v.gr., un macrófago), que lo fragmenta en péptidos pequeños. En la célula presentadora de antígeno ciertos fragmentos o epítopes son tomados por las moléculas clase II del complejo principal de histocompatibilidad (CPH) y transportados hacia la superficie de la CPA. El fragmento peptídico, que se localiza en el surco de la molécula del CPH, se fija entonces al RCT. La necesidad de que el antígeno sea presentado en asociación con una molécula del CPH se denomina restricción del CPH. Por lo tanto, los RCT no reconocen antígenos nativos, sino solo partes procesadas del mismo. Por el contrario, los receptores de las células B sí reconocen a los antígenos nativos.
Una característica de la respuesta inmunológica es su capacidad para incrementar el número de linfocitos antígeno-específicos después de un estímulo antigénico; por lo tanto, una exposición posterior al mismo antígeno proporciona una respuesta más rápida y de mayor intensidad. La base de esta respuesta aumentada es la proliferación de linfocitos antígeno-específicos y la producción de células de memoria despúes de la interacción con el antígeno.
Es importante que la respuesta inmunológica sea capaz de distinguir entre lo propio y lo que no es. En caso contrario, las células T y los anticuerpos atacarían en forma constante a células autólogas y componentes tisulares. Cuando por primera vez el Dr. Paul Ehrlich pensó acerca de esta posibilidad en 1901, afirmó que no podían suceder reacciones contra lo propio.1 Medio siglo después, Sir Frank Macfarlane Burnet postuló que el organismo debía poseer una manera de prevenir tales reacciones y sugirió que en el estado prenatal la interacción de los antígenos propios con los linfocitos antígeno-específicos provocaba la eliminación de estos linfocitos.2 El adulto sería, por lo tanto, incapaz de responder contra los antígenos propios, un estado que Burnet denominó tolerancia inmunológica. De hecho, es la necesidad para explicar la capacidad de la respuesta inmune para discriminar entre lo propio y lo no propio la que ha originado muchos de los conceptos actuales sobre el sistema inmunológico.
Autoinmunidad
Sin embargo, el estado de tolerancia inmunológica puede romperse y, en circunstancias anormales, pueden producirse anticuerpos y células sensibilizadas (antígeno-reactivas) que se dirijan contra antígenos propios. Este fenómeno se denomina autoinmunidad.
Mecanismos supresores
El concepto de tolerancia inmunológica se ha ampliado desde que Burnet lo postuló y ahora esta bien establecido que el sistema inmunológico puede responder a un antígeno, no sólo mediante la producción activa de varios agentes inmunológicos como serían los anticuerpos y las células sensibilizadas, sino también por medio de la activación de un número de mecanismos supresores diferentes que puedan disminuir estas manifestaciones. De hecho, la reacción final que se detecta después de la inyección de un antígeno es tal vez el resultado de un balance entre los mecanismos supresor y activador; por lo tanto, en ciertas situaciones clínicas, una respuesta inmunológica excesivamente severa puede ser debida a un defecto en los mecanismos supresores. Al contrario, en estados de inmunodeficiencia, la anormalidad puede no sólo deberse a la ausencia de un tipo particular de linfocitos, sino que también puede ser ocasionada por una respuesta supresora exagerada.
MECANISMOS SECUNDARIOS
Aunque una función del sistema inmunológico, la neutralización de toxinas producidas por microrganismos invasores, es llevada a cabo directamente por los anticuerpos producidos, muchas otras funciones, incluyendo la defensa del organismo frente a agentes infecciosos, el secuestro y remoción de material extraño y la destrucción de células que han sufrido una transformación maligna, son llevadas a cabo por mecanismos secundarios. Estos mecanismos se consideran secundarios no porque sean de menor importancia, sino porque son desencadenados por la interacción inicial entre el antígeno y el anticuerpo o los linfocitos T, y porque no son muy específicos. Sin embargo, los mecanismos secundarios pueden regular y amplificar la respuesta inmunológica. Los resultados de tales interacciones incluyen la producción de citocinas por linfocitos y otras células, la liberación de sustancias mediadoras a partir de las células cebadas y basófilos cubiertos con anticuerpos, la activación de las proteínas del complemento y la expresión de moléculas de integrina sobre las células. La alteración de la permeabilidad vascular, la expresión de receptores para leucocitos en las células endoteliales y la liberación de factores quimiotácticos, a través de estos mecanismos secundarios, atraen un gran número de células de otros tipos a la reacción. Estas células contribuyen en mucho a la inflamación resultante, ayudando en el proceso fagocítico y a la eliminación de antígenos extraños.
CONTROL GENETICO
La respuesta inmunológica está controlada por los genes de la respuesta inmune (Ir). Los genes Ir y los genes que controlan la producción de los antígenos del CPH (también conocidos como antígenos de histocompatibilidad o HLA en los humanos) se localizan muy cerca unos de otros en el mismo cromosoma (cromosoma 6). Los antígenos HLA identifican a las células autólogas como propias y las distinguen de las células de otros individuos. Los genes Ir y HLA constituyen parte del CPH, que tiene un papel crucial en la respuesta inmunológica. Las características del CPH explican por qué algunos individuos pueden no responder a ciertos antígenos. Por ejemplo, aunque los RCT reconocen epítopes unidos a las moléculas del CPH (ver antes), algunos péptidos antigénicos no se acomodan en el surco de una molécula de CPH específica de un individuo. Por lo tanto, la célula T adecuada no reaccionará al epítope y la persona no será capaz de montar una respuesta inmunológica contra él.
La capacidad del sistema inmunológico para discriminar entre tantas configuraciones moleculares diferentes es igualada en el campo de la biología sólo por la memoria y especificidad que tiene el cerebro. El sistema inmunológico puede recordar que ha tenido contacto previo con un antígeno y, por lo tanto, es capaz de responder con una mayor capacidad y amplificar aún más el proceso utilizando un cierto número de sistemas secundarios. En condiciones normales el sistema inmunológico puede distinguir entre antígenos extraños y propios, y la totalidad del proceso se encuentra bajo control genético.
Organos linfoides y circulación de linfocitos
El sistema inmune consta de diversos órganos linfoides, incluyendo el timo, los ganglios linfáticos, el bazo y las amígdalas, agregados de tejido linfático en órganos no linfoides, como las placas de Peyer en el intestino, grupos de células linfoides dispersas en todos los tejidos conjuntivos y epiteliales del organismo, así como en la médula ósea y la sangre, y una variedad de células individuales que circulan entre los diversos órganos linfoides y el resto del organismo. Aunque las células inmunológicamente activas de este sistema son los linfocitos, que se dividen en varios tipos, muchas otras células, incluyendo monocitos, macrófagos, granulocitos (neutrófilos, eosinófilos, basófilos y células cebadas) y plaquetas, desempeñan funciones accesorias importantes en el sistema inmunológico.
Los linfocitos derivan de células tronco (o células madre). En un principio éstas aparecen en el saco vitelino y en el hígado fetal; en la etapa tardía del desarrollo prenatal, algunas se originan de la médula ósea. Las células tronco se diferencían en linfocitos en los órganos linfoides primarios, que son básicamente el timo y la médula ósea.
ORGANOS LINFOIDES PRIMARIOS
Timo
El timo, que se origina en el embrión a partir de la tercera y cuarta bolsas faríngeas, se localiza en el mediastino anterior y está formado por múltiples lóbulos, cada uno de los cuales tiene una médula y una corteza [ver figura 3]. Los experimentos han indicado la importancia de este órgano al demostrar que los ratones en los que el timo se ha extirpado al nacimiento no tienen células T en sus órganos linfoides.3
Las células tronco de la médula ósea (destinadas a convertirse en células T) entran al timo, en donde proliferan y se diferencían. Al principio penetran a la corteza del timo, después migran hacia la médula y por último se movilizan hacia el sistema linfoide periférico. Los genes de las células T que entran al timo codifican RCT que tienen la capacidad de reaccionar con muchas moléculas diferentes del CPH, antígenos extraños y autoantígenos. A través de un proceso de selección positiva y negativa dentro del timo, más del 90 porciento de las células T mueren. Las que dejan el timo tienen restricción para el CPH. Las células T que reconocen autoantígenos son eliminadas o inactivadas.
Rearreglo de los genes del receptor de la célula T Los genes que codifican los RCT sufren rearreglos y las regiones V, de unión (J), de diversidad (D) y constante (C) se unen para constituir dos tipos de RCT, cada uno formado por dos cadenas. Cada tipo de RCT se asocia con una especificidad particular. Primero, los genes que codifican las cadenas g y d sufren rearreglos para formar el RCT-gd, después los genes que codifican las cadenas a y b se rearreglan para formar el RCT-ab. Después de ocurrida la combinación genética, el RCT se expresa en la superficie de la célula. Los RCT tienen aún más diversidad que las inmunoglobulinas porque tienen mayor número de genes para la región J y la región D, lo que permite codificar un repertorio preseleccionado de hasta 1015 especificidades. Al mismo tiempo que ocurre el rearreglo, se expresan varias moléculas accesorias importantes, incluyendo CD4, CD8, CD2 y CD34,5 [ver tabla 1]. (CD por las siglas en inglés de grupo de diferenciación). Relativamente pocas células T son CD4-, CD8- RCT-gd, cuya función no se conoce del todo. La mayoría se vuelven células RCT-ab que expresan en un principio CD4 y CD8 y que después, por selección positiva (ver adelante), se vuelven CD4+, CD8- RCT-ab (cooperadoras) o CD4-, CD8+ RCT-ab (que tienen funciones citotóxicas, supresoras o ambas). En todas las células T el CD3 se asocia con el RCT [ver figura 4].
Selección positiva La selección positiva está controlada por células epiteliales de la corteza tímica.6 Debido a que las células T pueden reaccionar a los antígenos solo en asociación con el CPH propio, solo las células T con un RCT que puede fijarse al CPH propio se seleccionan. Cuando estas células reaccionan con el CPH propio de las células epitaliales del timo, las células de tipo CD4+, CD8+ RCT-ab que se fijan a moléculas del CPH de clase II se convierten en CD4+ y CD8-, reduciendo la expresión del CD8 y aumentando la del complejo RCT-CD3. Por el contrario, las células CD4+, CD8+ que se fijan a moléculas del CPH de clase I disminuyen la expresión de CD4 y se vuelven células CD4-, CD8+ RCT-ab. De este modo se seleccionan las células CD4+ RCT-ab y las células CD8+ RCT-ab con restricción para el CPH propio. Se supone que las células T que se unen, aún en forma débil, a las moléculas del CPH con ayuda del ligando CD4 o CD8, serán capaces de unirse con más intensidad a las células presentadoras de antígeno, por lo que no se requieren antígenos extraños para la selección positiva. El mecanismo de la selección positiva ha sido confirmado por estudios en los que se eliminó la molécula del CPH clase II en ratones7 y por experimentos en los que se emplearon ratones transgénicos para el RCT.8 La mayoría de las células T no interactúan con el CPH propio y sufren muerte programada, o apoptosis.
Selección negativa El proceso de selección negativa elimina las células T que tienen un RCT con afinidad intensa a los autoantígenos. Si no se eliminaran, estas células podrían causar enfermedades autoinmunes. Existen muchos autoantígenos en las células epiteliales del timo, y los estudios han demostrado que ciertos autoantígenos pueden ser presentados por varias células presentadoras de antígenos en el timo, como macrófagos y células dendríticas. Es posible que la selección negativa para algunos autoantígenos ocurra cuando las células T se trasladan hacia el sistema linfoide periférico después de dejar el timo. La interacción de alta afinidad entre el autoantígeno presentado por la molécula del CPH y el RCT en las células T inmaduras puede desencadenar la muerte celular. Por el contrario, la interacción entre un antígeno presentado por la molécula del CPH y el RCT de una célula T madura ocasiona activación celular.
Migración de las células T hacia los órganos linfoides secundarios Dos a tres días después de que las células tronco entran en el timo, los linfocitos migran a través de la pared de las venas poscapilares de la médula. Entran al torrente sanguíneo y se localizan en los órganos periféricos del sistema linfoide. Una vez ahí, los linfocitos dejan el torrente sanguíneo, de nuevo a través de las vénulas poscapilares, y entran a las regiones timo dependientes del sistema linfoide periférico, a la corteza interna de los ganglios linfáticos, a las vainas periarteriales del bazo y a las zonas intranodulares de las placas de Peyer, las amígdalas y el apéndice. Sin embargo, parece ser que la mayoría de las células T del intestino no derivan del timo, sino que se diferencían in situ en el epitelio intestinal. Estas células son CD4-, CD8- RCT-ab o células CD4-, CD8- RCT-gd.9
Si al momento de nacer el timo está ausente, el número total de linfocitos en sangre es menor que el normal y hay una carencia notable o ausencia de linfocitos T. Las áreas dependientes del timo del sistema linfoide periférico también carecen de linfocitos. Este estado se relaciona con una afección notable de las funciones inmunológicas mediadas por células, como la capacidad para resistir ciertas infecciones y para rechazar tejidos transplantados. Está aumentada la susceptibilidad para ciertos tumores; también existe depresión en las respuestas con anticuerpos que requieren de cooperación de los linfocitos T. Algunos de los efectos de la timectomía en animales de experimentación pueden ser parcialmente corregidos con extractos de timo. Se han aislado varias hormonas de probable estirpe tímica, incluyendo la timosina, la timulina y la timopoyectina. Se considera que estas hormonas favorecen la maduración de las células T e, in vitro, facilitan ciertas funciones de las células T.
El timo involuciona con la edad, lo cual podría explicar el desarrollo de deficiencias en el sistema inmunológico que ocurre en personas ancianas.
Tejido equivalente al de la bolsa y médula ósea
Además del timo, las aves cuentan con otro órgano linfoide primario, la bolsa de Fabricio, que también se deriva del epitelio intestinal en el embrión y está situada cerca de la cloaca. Las células tronco entran en la bolsa de Fabricio, donde se diferencían en linfocitos B, capaces de producir anticuerpos humonales. Por medio de manipulación quirúrgica u hormonal puede evitarse el desarrollo de la bolsa. Los pollos en quienes se ha efectuado extipación de la bolsa tienen una deficiencia completa de células B (y de células plasmáticas) y, por lo tanto, son incapaces de producir anticuerpos humorales. Sin embargo, estos animales son capaces de rechazar injertos y muestran reacciones mediadas por células; en comparación con las aves timectomizadas, muestran sólo una disminución mínima en el número de linfocitos en sangre. Este hallazgo indicaría que la mayoría de sus linfocitos circulantes son células T.
En los mamíferos no parece existir un órgano que sea equivalente a la bolsa de Fabricio. En la actualidad las evidencias sugieren que las células tronco pueden diferenciarse a células B en la médula ósea y en los órganos linfoides periféricos.
ORGANOS LINFOIDES SECUNDARIOS
Ganglios linfáticos
Los delicados capilares linfáticos situados en la periferia de los ganglios linfáticos pueden ser atravesados con facilidad por varias células errantes en el tejido conjuntivo, así como por macromoléculas exógenas y endógenas [ver figura 7]. Estas células y macromoléculas son transportadas a los ganglios linfáticos a través de vías aferentes. Los sinusoides de los ganglios linfáticos contienen macrófagos que pueden fagocitar antígenos y partículas. Los linfocitos circulan a través de los sinusoides y salen de los ganglios linfáticos por las vías eferentes. Por los linfáticos eferentes salen del ganglio un número 50 veces mayor de células, la mayor parte linfocitos, de los que ingresan por las vías aferentes. ¿De dónde provienen estas células? Menos del cinco porciento se deriva de precursores situados en los propios ganglios linfáticos. La mayor parte proviene de la circulación sanguínea, hecho que destaca la importancia de la recirculación linfocítica. La mayor parte de estos linfocitos sanguíneos son linfocitos T de vida prolongada que entran en los ganglios linfáticos a través de vénulas poscapilares situadas en la profundidad de la corteza. Estas células circulan por la corteza y siguen por la médula y los linfáticos eferentes. Unos cuantos linfocitos B también provienen de la circulación sanguínea; éstos parecen pasar en forma más indiscriminada a todas las áreas de los ganglios linfáticos, aunque algunos se localizan en los centros germinales de la corteza.
La mayor parte de los linfocitos B situados en los ganglios linfáticos se encuentran en los centros germinales de la corteza periférica. Cuando los ganglios linfáticos han sido estimulados por antígenos capaces de inducir la formación de anticuerpos, los centros germinales aumentan de tamaño y, por lo tanto, contienen muchos linfoblastos. A diferencia de las células T, la mayor parte de las células B no tienden a emigrar.
Los complejos antígeno-anticuerpo son captados por los macrófagos y por las células dendríticas foliculares en los senos subcapsulares de los ganglios linfáticos. Estas células dendríticas poseen muchos receptores para los componentes C3 del complemento y para la porción Fc de la IgG. Pueden retener el antígeno no degradado en la forma de complejos antígeno-anticuerpo-C3 durante meses. Es probable que este antígeno atrapado proporcione el estímulo para la formación de los centros germinales, donde proliferan las células B de memoria. Además, las células dendríticas foliculares producen cuerpos cubiertos de complejos inmunes denominados icosomas. Estos icosomas están dispersos en los centros germinales, se unen a las células B y sufren endocitosis.10 Entonces el antígeno es procesado y presentado a las células T. También se encuentran algunas células T en los centros germinales, en donde, a través de la producción de citocinas, ayudan a las células B estimuladas por un antígeno a proliferar y diferenciarse.
Las áreas paracorticales de los ganglios linfáticos contienen células interdigitantes (también denominadas dentríticas). Estas células tienen antígenos de clase II del CPH sobre su superficie y presentan el antígeno a las células T. Se considera que algunas de estas células interdigitantes se derivan de las células de Langerhans que adquieren el antígeno mientras están en la piel y posteriormente emigran a los ganglios linfáticos.
La formación de compartimientos en los ganglios linfáticos (centros germinales con linfocitos B inmóviles, áreas paracorticales con linfocitos T que viajan con rapidez, sinusoides llenos de macrófagos y la malla reticular de células dendríticas que tiende a mantener los antígenos por tiempo prolongado) parece facilitar las interacciones entre los diferentes tipos de células que se requieren para generar una respuesta inmunológica.
La función de los ganglios linfáticos puede estar afectada de forma importante en estados de inmunodeficiencia. Por ejemplo, cuando el timo está ausente existe una pérdida notable de linfocitos en las áreas paracorticales. En la agamaglobulinemia ligada al X hay pérdida completa de linfocitos B en los folículos germinales de los ganglios linfáticos y en ningún sitio se observan células plasmáticas.
Bazo
En el bazo los linfocitos se concentran en la pulpa blanca que rodea a las arteriolas centrales, formando las llamadas vainas linfoides periarteriolares [ver figura 8]. Los linfocitos T se encuentran sobre todo en la región central de estas vainas linfoides, mientras que los linfocitos B localizados en los centros germinales están más concentrados hacia la periferia. La pulpa blanca está separada de la pulpa roja por una zona marginal. Los linfocitos entran y salen de las vainas linfoides periarteriolares a través de los capilares de las arteriolas centrales en la zona marginal. Algunas células plasmáticas inmaduras pasan de la zona marginal a la pulpa roja.
OTROS TEJIDOS LINFOIDES
Los linfocitos se encuentran también en otros lugares. El tejido linfoide relacionado con el intestino (TLRI) incluye las placas de Peyer y el apéndice. Estos tejidos contienen regiones con predominio de linfocitos T o B. En el intestino delgado también se encuentran muchos linfocitos, dentro de la lámina propia de la mucosa de las vellosidades y entre las células epiteliales de la mucosa. El tejido linfoide relacionado con las mucosas (TLRM) se encuentra en el aparato respiratorio, el intestino y el aparato urinario. Además, linfocitos aislados infiltran los tejidos epiteliales y conjuntivos de la mayor parte del organismo.
CIRCULACION DE LINFOCITOS
Existen tres tipos principales de circulación de los linfocitos: (1) la siembra de las células tronco provenientes del hígado fetal o de la médula ósea en los órganos linfoides primarios y la subsecuente diferenciación y distribución de estas células en el sistema linfoide periférico, (2) la recirculación sangre-linfa-sangre de linfocitos, y (3) la distribución de células efectoras a regiones particulares del organismo.
En el primer tipo de circulación, las células T tronco provenientes de la médula ósea penetran en el timo, donde proliferan y se diferencían en células inmunocompetentes en un plazo de dos a tres días; estas células son sembradas en los órganos linfoides periféricos. Además de las células T precursoras, la médula ósea también contiene algunas células T diferenciadas que son inmunocompetentes. La presencia de células inmunocompetentes en la médula ósea se demuestra en las notables reacciones de injerto contra huésped, mediadas por células T, que se presentan cuando se emplea médula ósea histoincompatible en la realización de trasplantes. Las células B tronco se movilizan de la médula ósea a los órganos linfoides secundarios. Los linfocitos B y T maduros no vuelven a entrar a los órganos linfoides primarios en cantidades significativas.
En el segundo patrón, la recirculación de linfocitos de la sangre, a través de los ganglios hacia los vasos linfáticos y de regreso a la sangre, es mucho más rápida y no entraña proliferación celular. Los tiempos promedio de tránsito de los linfocitos pequeños, la mayoría de los cuales son T con vida prolongada, son de 0.6 horas en la sangre, cinco a seis horas en el bazo, y 15 a 20 horas en los ganglios linfáticos. El tiempo de tránsito para los linfocitos B es más largo; por ejemplo, las células B pasan 30 horas en los ganglios linfáticos. Los linfocitos entran a los ganglios linfáticos a través de una zona especializada de la vénula poscapilar formada por células endoteliales altas y metabólicamente activas. Estas vénulas modificadas son denominadas vénulas endoteliales altas (VEA). Las citocinas producidas por las células T activadas cambian las vénulas planas por vénulas endoteliales altas. Los linfocitos se unen mejor a éstas últimas y después de unirse son capaces de viajar a través de las mismas.11
Las células T permanecen en la porción interna de la corteza, mientras que las B se mueven con lentitud hacia afuera, hasta los centros germinales. Se supone que en alguna parte de su trayecto las células B deben atravesar las áreas dependientes del timo y dirigirse hacia la médula, debido a que es ahí donde se encuentran la mayoría de las células plasmáticas (que se derivan de los linfocitos B). Si los linfocitos son extraídos del conducto tórácico sin permitir su reentrada a la sangre, las zonas dependientes del timo del sistema linfoide periférico se agotan de linfocitos mucho más rápidamente que los centros germinales; esta disminución selectiva es otra indicación de que las células B en estos órganos se mueven con más dificultad que las células T.
El tercer patrón de circulación de linfocitos es el movimiento de las células efectoras o de las células de memoria hacia los sitios de inflamación y hacia las mucosas. Las células T con marcador de superficie CD45RA se fijan en forma preferencial a las vélulas endoteliales altas de los ganglios linfáticos; por el contrario, las células T de memoria con el marcador de superficie CD45RO viajan de preferencia a la piel, las mucosas y otros tejidos normales, así como a sitios de inflamación. Los linfocitos tienen diversos receptores de superficie que influyen en el sitio al que se dirigen o donde se alojan. Estos receptores incluyen la L-selectina, el antígeno muy tardío-4 (VLA-4) y el antígeno asociado a la función del linfocito-1 (LFA-1). Las vénulas endoteliales altas de varios tejidos tienen también en su superficie moléculas de adhesión que contienen ligandos endoteliales selectivos para los tejidos, llamados adresinas vasculares. Estas adresinas, como la E-selectina, la molécula de adhesión intercelular-1 (ICAM-1), la ICAM-2, y la molécula de adhesión a células vasculares-1 (VCAM-1), se fijan a receptores específicos en los linfocitos. La expresión de estas adresinas es inducida por diversas citocinas.
Células inmunocompetentes de la respuesta inmunológica
Los linfocitos, las células principales en la respuesta inmunológica y las únicas células que reconocen antígenos específicos, constituyen una familia de células que tienen un núcleo redondo de localización central, carecen de gránulos específicos y tienen un citoplasma basófilo que contiene ribosomas libres. Las células T regulan la respuesta inmunológica, participan en las reacciones mediadas por células e inducen a los linfocitos B para producir anticuerpos. Las células B se diferencían en células plasmáticas productoras de anticuerpos. Ni los linfocitos T ni los B constituyen una población celular homogénea; cada grupo incluye diferentes subgrupos que pueden distinguirse por sus marcadores de superficie y por su función. Además, existe un grupo heterogéneo de linfocitos que no son ni células T ni B. Mientras que algunas de estas células pueden ser precursoras de las células T o B, otras parecen formar otro grupo o grupos generales. Estos grupos o subgrupos de linfocitos no pueden diferenciarse unos de otros por técnicas morfológicas. La fijación de anticuerpos monoclonales a marcadores de superficie es en la actualidad la técnica más específica para distinguir entre los subgrupos de linfocitos.
CELULAS T
La observación de que algunos linfocitos humanos pueden fijar eritrocitos de carnero, formando rosetas, llevó a la designación del marcador original de los linfocitos T humanos. El uso de anticuerpos monoclonales reveló que los eritrocitos de carnero se fijan en una glucoproteína de 55 kd situada sobre la superficie del linfocito, en la actualidad este antígeno se denomina CD2. Todas las células T maduras y los timocitos medulares expresan el antígeno CD2, así como otro antígeno de superficie, conocido como CD3. Los linfocitos portadores de CD2 están ausentes o muy disminuidos en la sangre de los niños con aplasia tímica, pero aparecen con rapidez después de colocar un injerto de timo.
Subgrupos de células T
Se han identificado varios subgrupos de células T que tienen diferentes funciones y se les ha analizado empleando una serie de anticuerpos monoclonales dirigidos contra las moléculas de superficie.12 Del 50 al 65 porciento de las células T periféricas muestran una proteína de superficie de 57 kd conocida como CD4, la cual se reconoce por los anticuerpos monoclonales OKT4 y anti-Leu-3. Del 25 al 35 porciento de las células T periféricas muestran una glucoproteína de 32 kd llamada CD8, que se reconoce por los anticuerpos monoclonales OKT8 y anti-Leu-2. Aunque CD4 y CD8 se expresan juntas en los timocitos en la etapa II, sólo una u otra se expresa en los subgrupos complementarios de las células T maduras (células T CD4+ y CD8+). Las células T CD4+ reconocen antígenos cuando se presentan asociados con las moléculas de clase II del CPH (antígenos HLA-D); las células T CD8+ reconocen antígenos en la vecindad de moléculas de clase I del CPH (antígenos HLA-A, HLA-B y HLA-C).
Las moléculas CD4 y CD8 se acoplan a una tirosina cinasa específica denominada p56lck en el citoplasma de la célula T.12 Esta cinasa se relaciona con la familia del protooncogen src que está implicado en la transformación y activación de algunas células de mamíferos, y cinasas similares forman parte de la cola citoplasmica de receptores para hormonas de crecimiento, como el factor de crecimiento epidémico, el factor de crecimiento derivado de las plaquetas y la insulina. En estudios in vitro, los complejos CD4-p56lck y CD8-p56lck fosforilaron la porción citoplásmica de las subunidades g, d, e, y en especial la subunidad s de CD3, pero no fosforilaron otros 12 antígenos CD probados.12 Esta interacción explica cómo CD4 y CD8 interactúan con el complejo CD3-RCT y pueden desempeñar un papel importante en la activación de la célula T. El papel crucial de la derivación de CD4-RCT en la señal para la activación de la célula T se apoya en otros dos estudios. Es interesante que el antígeno común leucocitario CD45 parece tener un dominio intracelular con actividad de fosfatasa de tirosina proteica y podría regular la activación de las células T por la cinasa p56lck.
Las células T CD4+, pueden diferenciarse en células TH1 y TH2 según las linfocinas que producen.13,14 Las células TH1 producen interleucina-2 (IL-2) e interferón gama, que son importantes en las reacciones inmunológicas mediadas por células. Las células Th2, que producen interleucina-4 (IL-4), interleucina-5 (IL-5), interleucina-6 (IL-6) e interleucina-10 (IL-10), ayudan a las células B en la producción de anticuerpos. Las linfocinas que son producidas por cada uno de estos tipos celulares influyen en el otro tipo celular. El interferón gama producido por las células TH1 puede inhibir la función de las células TH2, mientras que la IL-10, que es producida por células TH2 (así como por monocitos, macrófagos y células B), puede inhibir la función de las células TH1. La IL-10 no actúa en forma directa sobre los TH1 sino sobre células accesorias de las que éstos dependen. Las citocinas formadas por otras células también pueden influir en las subpoblaciones de linfocitos T. Por ejemplo, la interleucina-12 (IL-12) favorece la diferenciación de las células TH1.
CELULAS B
Los linfocitos B son las células productoras de inmunoglobulinas del sistema inmune, y pueden ser identificadas por la presencia de inmunoglobulinas en su superficie. Estas células con receptores para inmunoglobulinas positivos en la superficie de su membrana (SmIg+) y constituyen del 5 al 15 porciento de los linfocitos en sangre periférica [ver figura 11]. Esta definición no incluye a los precursores de las células B, que tienen IgM en su citoplasma pero no en su superficie, ni a las células plasmáticas SmIg- que se originan a partir de las células B. La mayor parte de las células B tienen IgM e IgD en su superficie y cerca de la cuarta parte del total de estas células sólo poseen inmunoglobulinas de alguna de estas dos clases. Uno porciento de las células B muestra IgG o IgA Los linfocitos T y B constituyen del 80 al 95 porciento de los linfocitos en sangre periférica. El resto de las los linfocitos presentes en sangre periférica constituyen una población heterogénea de células (ver adelante).
Sobre la superficie de las células B se encuentra un receptor del complemento llamado receptor 2 del complemento (RC2), el cual se une a los factores C3d, C3dg y al factor iC3b. Las células B también poseen receptores para el virus de Epstein-Barr y para la porción Fc de la IgG. La molécula CD20, una fosfoproteína de 35 kd que se encuentra sobre la superficie de las células B, es un marcador de los linfocitos B utilizado con frecuencia. Algunos aloantígenos están presentes sobre la superficie de los linfocitos B, pero no sobre la superficie de los linfocitos T. Estos antígenos se detectan usando sueros de mujeres embarazadas y algunos antisueros para antígenos purificados obtenidos de las líneas celulares de linfoblastos B. Algunos precursores de las células B carecen del receptor SmIg, pero poseen estos aloantígenos.
Un grupo de células B, las células de memoria, aumentan la respuesta inmune a los antígenos ya conocidos, y constituyen la llamada respuesta inmune secundaria, respuesta anamnésica o respuesta de refuerzo. Estas células B de memoria sufren mutaciones somáticas en las regiones variables de sus genes de inmunoglobulina. Cuando esta mutación somática ocurre en los centros germinales de los ganglios linfáticos que contienen antígenos unidos a las células dendríticas foliculares, se presenta selección de las células B de memoria que tienen receptores de alta afinidad para los antígenos.
Un subgrupo de células B, los llamados linfocitos B1, se desarrollan en fases tempranas y tienen vida media muy prolongada. Los experimentos realizados sobre la capacidad de los progenitores de células B para reconstituir diferentes poblaciones de linfocitos B en ratones adultos radiados sugieren que las células B1 y los linfocitos B regulares se producen a través de linajes diferentes.15 Se encuentran progenitores de células B1 en el higado fetal y en el epiplón embrionario, pero no en la médula ósea adulta. Las células B1 que expresan CD5 en su superficie se conocen como B1a, y las que no se denominan células B1b. Las células B1 se asocian con frecuencia con producción de autoanticuerpos. También elaboran grandes cantidades de IL-10.
Al hacer el recuento de las células SmIg+ es importante recordar que las inmunoglobulinas producidas por las células B pueden fijarse de manera pasiva a la superficie de las membranas de células no-B. Por ejemplo, las células que tienen receptores Fc fijan complejos de inmunoglobulina; más aún, los anticuerpos fluorescentes para inmunoglobulinas, que se emplean con frecuencia para identificar a las células SmIg+, también pueden fijarse a los receptores Fc. Este problema puede evitarse empleando anticuerpos fluorescentes F(ab')2, esto es, la inmunoglobulina cuya fracción Fc ha sido eliminada. Además, la presencia de autoanticuerpos contra linfocitos T incrementará el número de células SmIg+ que son confundidas con células B. Estos anticuerpos antilinfocíticos se han encontrado en varias enfermedades, incluyendo el lupus eritematoso generalizado (LEG), la mononucleosis infecciosa, la neumonía atípica, la artritis reumatoide y varios trastornos linfoproliferativos. La presencia de estos anticuerpos se sospecha por lo general cuando los porcentajes de células CD3+ y células SmIg+ suman más del 100 porciento. Las inmunoglobulinas absorbidas de manera pasiva pueden eliminarse incubando las células toda una noche en ausencia de suero humano o por medio de un ligero tratamiento con tripsina. En este último caso las células se cultivan poco tiempo después del tratamiento con tripsina para que las células B genuinas puedan regenerar su SmIg.
CELULAS ASESINAS NATURALES
Las células asesinas naturales (NK, por sus siglas en inglés, n. del t.) son linfocitos granulares de gran tamaño que no poseen el complejo RCT-CD3 característico de las células T ni el SmIg de las células B. In vitro, estas células pueden destruir diversas células tumorales o infectadas por virus de modo no específico. Esto es, no requieren de una sensibilización previa o de la presencia de anticuerpos para ser citotóxicas. Sus gránulos contienen proteínas formadoras de poros que pueden mediar la lisis celular. Las células NK no tienen memoria ni muestran restricción de CPH. No se conoce con precisión al precursor de las células NK, pero se piensa que es diferente a la célula tronco tímica (precursora de los linfocitos T) y a la célula pro-B (precursora de los linfocitos B).
Las células asesinas naturales poseen varios marcadores de superficie, incluyendo el CD56, una glucoproteína de 140 kd también denominada NKH1, el CD2, que también está presente en las células T y el CD16, un receptor Fc. El CD16 se asocia con la cadena s del complejo RCT-CD3. Esta cadena participa en la transducción de señales desencadenada por los anticuerpos que se fijan al CD16, lo que activa la célula.
La IL-12 estimula la proliferación de las células NK y su producción de interferón gama, que es importante en muchas reacciones inmunológicas.16
Una población de células T citotóxicas no restringida al CPH puede lisar algunas de las mismas células blanco que destruyen las células asesinas naturales. Estas células T citotóxicas tienen un marcador de superficie que puede detectarse por medio del anti-CD56, pero no por anti-CD16. Además, expresan el complejo RCT-CD3.17
Una pequeña proporción de las células nulas de la sangre son precursores mieloides y precursores de los linfocitos T y B inmaduros.
CELULAS PLASMATICAS
Por la influencia de los antígenos, los linfocitos T y células accesorias, las células B se diferencian en células plasmáticas, que son células maduras productoras de anticuerpos. Estas células son de mayor tamaño que los linfocitos y se caracterizan por un núcleo redondo y excéntrico con heterocromatina gruesa distribuida en un patrón en rueda de carreta. Las células plasmáticas tienen un citoplasma altamente basófilo, y un retículo endoplásmico bien desarrollado, con frecuencia organizado en capas concéntricas. Las células plasmáticas pueden estar repletas de material granular, que corresponde al anticuerpo que están produciendo [ver figura 12]. Algunas veces, una o más de las cisternas del retículo endoplásmico se encuentran distendidas por grandes inclusiones llamadas cuerpos de Russell; éstos son agregados de moléculas de inmunoglobulinas formadas de manera incompleta. Las células plasmáticas no tienen inmunoglobulinas en su superficie. Estas células son terminales, lo que significa que no se dividen. Los precursores inmaduros de las células plasmáticas, los plasmoblastos, son difíciles de distinguir de los linfoblastos y de los linfocitos grandes. En condiciones normales no se encuentran células plasmáticas en sangre periférica.
MONOCITOS Y MACROFAGOS
Los monocitos pertenecen al sistema fagocítico mononuclear, antes llamado sistema reticuloendotelial. Son grandes células mononucleares que constituyen del tres al ocho porciento de los leucocitos en sangre periférica. Su citoplasma es mucho más abundante que el de los linfocitos. Su núcleo es a menudo excéntrico y oval o reniforme [ver figura 13]. Los lisosomas llenos de enzimas degradantes aparecen como pequeñas vacuolas en el citoplasma. Los monocitos se originan a partir de los promonocitos, los cuales son precursores con una tasa elevada de división y que se encuentran en la médula ósea. Cuando las células maduras entran a la sangre periférica son llamadas monocitos; cuando abandonan la sangre e infiltran los tejidos, sufren cambios adicionales y entonces son conocidas como macrófagos.
Los macrófagos desempeñan un papel importante en la inducción de la respuesta inmunológica al ser uno de los tipos de células que presenta el antígeno a los linfocitos. También actúan como células efectoras, atacando ciertos microrganismos y células neoplásicas, y eliminando material extraño.
Los macrófagos contienen receptores específicos para anticuerpos y complemento, lo que incrementa su capacidad para fagocitar microrganismos cubiertos con estas moléculas. Los receptores de anticuerpos son para las porciones Fc de la IgG1 y de la IgG3; existe también un receptor Fc para la IgE. Tienen dos receptores del complemento: el receptor 1 (RC1) y el receptor 3 (RC3). El RC1 tiene alta afinidad por el componente C3b del complemento y menor afinidad por iC3b y C4b. El RC3, también llamado MAC-1, interactúa con iC3b y con ciertas moléculas de carbohidratos, incluyendo los antígenos que contienen carbohidratos del protozoario Leishmania.
Los macrófagos poseen también receptores para citocinas; el interferón gama es la principal citocina activadora de estas células.
Una pequeña proporción de los antígenos de clase II del CPH está presente en los monocitos; la expresión de las moléculas de clase II del CPH se incrementa considerablemente cuando se activan los macrófagos, lo cual puede ocurrir por varias linfocinas, incluyendo el interferón gama, el factor estimulador de colonias de granulocitos y macrófagos (FEC-GM), el factor activador de los macrófagos (FAM) y el factor inhibidor de la migración (FIM). Linfocinas como la interleucina-4 y el factor-b transformador del crecimiento (FCT-b) antagonizan esta activación. El antígeno Mo3e, una proteína de 50 kd, es un receptor de urocinasa. Este antígeno ha sido identificado por medio de anticuerpos monoclonales y parece ser específico de los macrófagos activados. El Mo3e no se ha encontrado en otras células hematopoyéticas.18
Los macrófagos producen un gran número de sustancias solubles que son importantes en la respuesta inmunológica y en el proceso de inflamación.19 Estas sustancias incluyen enzimas como el factor activador del plasminógeno y la elastasa, factores de crecimiento como el FEC-GM, citocinas como la interleucina-1 (IL-1), IL-6, IL-10, IL-12 y el factor de necrosis tumoral-a (FNT-a), factores que son críticos para combatir a los microrganismos, como los metabolitos del oxígeno y el óxido nítrico, componentes del complemento de la vía clásica y alterna, proteínas inflamatorias de macrófagos (PIM), y factores que facilitan la reparación tisular, como el factor de crecimiento de fibroblastos (FCF).
GRANULOCITOS
Neutrófilos
El granulocito predominante en la sangre es el neutrófilo. Se origina a partir de precursores en la médula ósea, su vida media en sangre es de cuatro a ocho horas, pero en los tejidos vive hasta tres veces más. Los neutrófilos son atraídos de la sangre hacia los lugares en donde hay inflamación por diversos factores quimiotácticos, incluyendo aquellos generados por activación de la cascada del complemento y algunos producidos por linfocitos estimulados por antígenos. Los neutrófilos tienen una importancia primaria en la defensa del organismo debido a su capacidad para fagocitar y destruir microrganismos. Este proceso es facilitado considerablemente por el complemento y los anticuerpos. La importancia de la fagocitosis por neutrófilos se ilustra de manera especial por el hecho de que en pacientes con deficiencia o ausencia total de estas células las infecciones piógenas ponen en peligro la vida.
Los neutrófilos también contienen gránulos lisosomales llenos de una variedad de enzimas degradantes y de pequeñas proteínas que pueden causar inflamación cuando son liberadas por las células. Muchos neutrófilos se encuentran en los lugares donde se localizan ciertas reacciones inmunopatológicas, en especial aquellas desencadenadas por complejos inmunes. Estas células están dotadas con receptores para IgG y para el componente C3b del complemento.
Eosinófilos
Los eosinófilos se caracterizan por poseer grandes gránulos rojos, que contienen proteínas básicas y muchas enzimas degradantes. La función exacta de los eosinófilos ha sido desde hace mucho tiempo un misterio, aunque se han sugerido varias funciones diferentes. Por ejemplo, se han encontrado muchos eosinófilos en tejidos donde se presentan reacciones inmunológicas que han sido desencadenadas por anticuerpos IgE, como en los pólipos nasales o en la pared bronquiolar de algunos enfermos con asma. Los eosinófilos son activados cuando el antígeno estimula a las células cebadas cubiertas con IgE para liberar un factor quimiotáctico de la anafilaxia para eosinófilos. El hallazgo de que los eosinófilos contienen diversas enzimas que pueden degradar a los mediadores de la hipersensibilidad inmediata, como la histamina, la sustancia de rección lenta de la anafilaxia (leucotrienos) y el factor agregante plaquetario, sugiere que dichas células pueden controlar o disminuir tales reacciones de hipersensibilidad. Los eosinófilos también son atraídos por los complejos antígeno-anticuerpo y pueden fagocitarlos. Algunos eosinófilos tienen receptores para Fc y C3b lo que les permite unirse a los complejos antígeno-anticuerpo o a los organismos que han desencadenado la vía alterna del complemento y están cubiertos con C3b.
Los eosinófilos se acumulan con frecuencia en sitios de infecciones producidas por helmintos. En la esquistosomiasis, por ejemplo, los eosinófilos se encuentran alrededor de las larvas y en los granulomas inducidos por los huevos. Debido a que los eosinófilos modulan la citotoxicidad mediada por células dependientes de anticuerpos (CCDA), se considera que participan en la resistencia a algunas infecciones parasitarias. Los eosinófilos pueden también contribuir a la inflamación y a cambios tisulares como los observados en pacientes con fibrosis endomiocárdica que se desarrolla después de hipereosinofilia prolongada. La capacidad de los eosinófilos para destruir larvas cubiertas con anticuerpos está aumentada por el factor reforzador citotóxico eosinofílico derivado de los monocitos, así como por el FNT-a y el FEC-GM. La liberación de estas citocinas por los macrófagos, células cebadas o linfocitos probablemente aumenta la inflamación producida por los eosinófilos. La IL-5 es crucial en la produción de eosinofilia; en los ratones, los anticuerpos anti-IL-5 disminuyen notablemente el número de eosinófilos en la sangre y los tejidos.
Basófilos y células cebadas
Los basófilos y las células cebadas tienen gránulos citoplásmicos metacromáticos. La estructura de los gránulos está formada por heparina unida a diversas aminas vasoactivas y otros mediadores. Los basófilos tienen un núcleo multilobulado y relativamente pocos gránulos metacromáticos, de gran tamaño, mientras que las células cebadas tienen un núcleo unilobular y gránulos citoplásmicos pequeños y más numerosos. Los basófilos se encuentran sobre todo en la sangre y en sitios de inflamación, mientras que las células cebadas están presentes principalmente en los tejidos. Aunque las dos células tienen funciones en común, se originan de diferentes precursores. Las células cebadas tienen más enzimas en sus gránulos que los basófilos y pueden liberar varias citocinas, incluyendo interleucina-3 (IL-3), IL-4, IL-5, IL-6 y FEC-GM. Ambos tipos de células tienen receptores Fc para IgE e IgG; de hecho, es la combinación del antígeno con la IgE ligada a estas células la responsable de la liberación de los mediadores en las reaciones de hipersensibilidad inmediata. Se ha demostrado que las células cebadas también poseen receptores para C3b.
Además de su papel en las reacciones de hipersensibilidad inmediata, los basófilos se encuentran en gran número en algunas reacciones tardías mediadas por células, como la hipersensibilidad por contacto en humanos, el rechazo a tumores o a injertos de piel y la hipersensibilidad a ciertos microrganismos en animales.
PLAQUETAS
La principal función de las plaquetas es participar en la coagulación sanguínea, pero también están implicadas en algunos de los mecanismos secundarios iniciados por la respuesta inmune. Las plaquetas poseen receptores Fc para la IgG e IgE y receptores para las moléculas de clase I del CPH sobre su superficie. Se adhieren y se agregan sobre las superficies endoteliales dañadas, liberando factores que producen aumento de la permeabilidad capilar y activan diversos componentes del complemento que en forma secundaria atraen leucocitos al lugar de la lesión. Bajo ciertas circunstancias se ha demostrado que las plaquetas participan en la defensa del huésped contra microrganismos invasores.
Bibliografía
DR. JOHN DAVID
Características del sistema inmune
Por la capacidad de distinguir lo propio de lo que no lo es, el sistema inmune es el mediador de la relación del individuo con el medio ambiente que le rodea. Cuando el sistema inmune funciona de manera apropiada, protege al organismo de las infecciones; cuando no, los fallos del sistema inmune producen algunas de las enfermedades más graves y difíciles que el médico puede encontrar en la práctica médica.
MEMORIA Y ESPECIFICIDAD INMUNOLOGICA
La experiencia con las infecciones ha demostrado que el organismo tiene memoria para enfermedades tales como la viruela, el sarampión o la parotiditis. Una vez que un individuo normal ha padecido una enfermedad como las mencionadas, el sistema inmune la reconoce y previene su recurrencia. Además de memoria, el sistema tiene especificidad, esto es, tiene la extraordinaria capacidad para discriminar entre diferentes antígenos, aun cuando sus estructuras estén estrechamente relacionadas entre sí.
Existen dos grupos principales de linfocitos, los linfocitos T (también llamado linfocitos dependientes del timo o células T) y los linfocitos B (también conocidos como linfocitos derivados de la médula ósea o células B). La capacidad de los linfocitos T y de los anticuerpos producidos por los linfocitos B para discriminar entre los antígenos está determinada por los genes de la región variable (V). Durante la maduración de los linfocitos T y B ocurre rearreglo de las secuencias de ADN en estos genes de la región V, además de mutaciones somáticas adicionales en las células B. Los varios cientos de genes de la región V que se forman codifican las inmunoglobulinas (anticuerpos) y los receptores de las células T (RCT), que pueden distinguir alrededor de un billón de diferentes especificidades antigénicas.
Existe un número enorme de linfocitos, cada uno de los cuales tiene un receptor de superficie que reconoce un solo determinante antigénico o epítope. Después de interactuar con un antígeno, los linfocitos T proliferan y se diferencían en células de memoria y en células que regulan la producción de anticuerpos por los linfocitos B y las respuestas efectoras mediadas por células de otros linfocitos T. Después de interactuar con un antígeno, los linfocitos B proliferan y se diferencían en células de memoria y células plasmáticas, encargadas de la producción de anticuerpos.
Antes de ser reconocido por las células T, el antígeno es captado por una célula presentadora de antígeno (CPA) (v.gr., un macrófago), que lo fragmenta en péptidos pequeños. En la célula presentadora de antígeno ciertos fragmentos o epítopes son tomados por las moléculas clase II del complejo principal de histocompatibilidad (CPH) y transportados hacia la superficie de la CPA. El fragmento peptídico, que se localiza en el surco de la molécula del CPH, se fija entonces al RCT. La necesidad de que el antígeno sea presentado en asociación con una molécula del CPH se denomina restricción del CPH. Por lo tanto, los RCT no reconocen antígenos nativos, sino solo partes procesadas del mismo. Por el contrario, los receptores de las células B sí reconocen a los antígenos nativos.
Una característica de la respuesta inmunológica es su capacidad para incrementar el número de linfocitos antígeno-específicos después de un estímulo antigénico; por lo tanto, una exposición posterior al mismo antígeno proporciona una respuesta más rápida y de mayor intensidad. La base de esta respuesta aumentada es la proliferación de linfocitos antígeno-específicos y la producción de células de memoria despúes de la interacción con el antígeno.
Es importante que la respuesta inmunológica sea capaz de distinguir entre lo propio y lo que no es. En caso contrario, las células T y los anticuerpos atacarían en forma constante a células autólogas y componentes tisulares. Cuando por primera vez el Dr. Paul Ehrlich pensó acerca de esta posibilidad en 1901, afirmó que no podían suceder reacciones contra lo propio.1 Medio siglo después, Sir Frank Macfarlane Burnet postuló que el organismo debía poseer una manera de prevenir tales reacciones y sugirió que en el estado prenatal la interacción de los antígenos propios con los linfocitos antígeno-específicos provocaba la eliminación de estos linfocitos.2 El adulto sería, por lo tanto, incapaz de responder contra los antígenos propios, un estado que Burnet denominó tolerancia inmunológica. De hecho, es la necesidad para explicar la capacidad de la respuesta inmune para discriminar entre lo propio y lo no propio la que ha originado muchos de los conceptos actuales sobre el sistema inmunológico.
Autoinmunidad
Sin embargo, el estado de tolerancia inmunológica puede romperse y, en circunstancias anormales, pueden producirse anticuerpos y células sensibilizadas (antígeno-reactivas) que se dirijan contra antígenos propios. Este fenómeno se denomina autoinmunidad.
Mecanismos supresores
El concepto de tolerancia inmunológica se ha ampliado desde que Burnet lo postuló y ahora esta bien establecido que el sistema inmunológico puede responder a un antígeno, no sólo mediante la producción activa de varios agentes inmunológicos como serían los anticuerpos y las células sensibilizadas, sino también por medio de la activación de un número de mecanismos supresores diferentes que puedan disminuir estas manifestaciones. De hecho, la reacción final que se detecta después de la inyección de un antígeno es tal vez el resultado de un balance entre los mecanismos supresor y activador; por lo tanto, en ciertas situaciones clínicas, una respuesta inmunológica excesivamente severa puede ser debida a un defecto en los mecanismos supresores. Al contrario, en estados de inmunodeficiencia, la anormalidad puede no sólo deberse a la ausencia de un tipo particular de linfocitos, sino que también puede ser ocasionada por una respuesta supresora exagerada.
MECANISMOS SECUNDARIOS
Aunque una función del sistema inmunológico, la neutralización de toxinas producidas por microrganismos invasores, es llevada a cabo directamente por los anticuerpos producidos, muchas otras funciones, incluyendo la defensa del organismo frente a agentes infecciosos, el secuestro y remoción de material extraño y la destrucción de células que han sufrido una transformación maligna, son llevadas a cabo por mecanismos secundarios. Estos mecanismos se consideran secundarios no porque sean de menor importancia, sino porque son desencadenados por la interacción inicial entre el antígeno y el anticuerpo o los linfocitos T, y porque no son muy específicos. Sin embargo, los mecanismos secundarios pueden regular y amplificar la respuesta inmunológica. Los resultados de tales interacciones incluyen la producción de citocinas por linfocitos y otras células, la liberación de sustancias mediadoras a partir de las células cebadas y basófilos cubiertos con anticuerpos, la activación de las proteínas del complemento y la expresión de moléculas de integrina sobre las células. La alteración de la permeabilidad vascular, la expresión de receptores para leucocitos en las células endoteliales y la liberación de factores quimiotácticos, a través de estos mecanismos secundarios, atraen un gran número de células de otros tipos a la reacción. Estas células contribuyen en mucho a la inflamación resultante, ayudando en el proceso fagocítico y a la eliminación de antígenos extraños.
CONTROL GENETICO
La respuesta inmunológica está controlada por los genes de la respuesta inmune (Ir). Los genes Ir y los genes que controlan la producción de los antígenos del CPH (también conocidos como antígenos de histocompatibilidad o HLA en los humanos) se localizan muy cerca unos de otros en el mismo cromosoma (cromosoma 6). Los antígenos HLA identifican a las células autólogas como propias y las distinguen de las células de otros individuos. Los genes Ir y HLA constituyen parte del CPH, que tiene un papel crucial en la respuesta inmunológica. Las características del CPH explican por qué algunos individuos pueden no responder a ciertos antígenos. Por ejemplo, aunque los RCT reconocen epítopes unidos a las moléculas del CPH (ver antes), algunos péptidos antigénicos no se acomodan en el surco de una molécula de CPH específica de un individuo. Por lo tanto, la célula T adecuada no reaccionará al epítope y la persona no será capaz de montar una respuesta inmunológica contra él.
La capacidad del sistema inmunológico para discriminar entre tantas configuraciones moleculares diferentes es igualada en el campo de la biología sólo por la memoria y especificidad que tiene el cerebro. El sistema inmunológico puede recordar que ha tenido contacto previo con un antígeno y, por lo tanto, es capaz de responder con una mayor capacidad y amplificar aún más el proceso utilizando un cierto número de sistemas secundarios. En condiciones normales el sistema inmunológico puede distinguir entre antígenos extraños y propios, y la totalidad del proceso se encuentra bajo control genético.
Organos linfoides y circulación de linfocitos
El sistema inmune consta de diversos órganos linfoides, incluyendo el timo, los ganglios linfáticos, el bazo y las amígdalas, agregados de tejido linfático en órganos no linfoides, como las placas de Peyer en el intestino, grupos de células linfoides dispersas en todos los tejidos conjuntivos y epiteliales del organismo, así como en la médula ósea y la sangre, y una variedad de células individuales que circulan entre los diversos órganos linfoides y el resto del organismo. Aunque las células inmunológicamente activas de este sistema son los linfocitos, que se dividen en varios tipos, muchas otras células, incluyendo monocitos, macrófagos, granulocitos (neutrófilos, eosinófilos, basófilos y células cebadas) y plaquetas, desempeñan funciones accesorias importantes en el sistema inmunológico.
Los linfocitos derivan de células tronco (o células madre). En un principio éstas aparecen en el saco vitelino y en el hígado fetal; en la etapa tardía del desarrollo prenatal, algunas se originan de la médula ósea. Las células tronco se diferencían en linfocitos en los órganos linfoides primarios, que son básicamente el timo y la médula ósea.
ORGANOS LINFOIDES PRIMARIOS
Timo
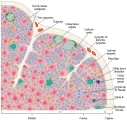
El timo, que se origina en el embrión a partir de la tercera y cuarta bolsas faríngeas, se localiza en el mediastino anterior y está formado por múltiples lóbulos, cada uno de los cuales tiene una médula y una corteza [ver figura 3]. Los experimentos han indicado la importancia de este órgano al demostrar que los ratones en los que el timo se ha extirpado al nacimiento no tienen células T en sus órganos linfoides.3
|
| Figura 3 |
| Estructura del timo |
Las células tronco de la médula ósea (destinadas a convertirse en células T) entran al timo, en donde proliferan y se diferencían. Al principio penetran a la corteza del timo, después migran hacia la médula y por último se movilizan hacia el sistema linfoide periférico. Los genes de las células T que entran al timo codifican RCT que tienen la capacidad de reaccionar con muchas moléculas diferentes del CPH, antígenos extraños y autoantígenos. A través de un proceso de selección positiva y negativa dentro del timo, más del 90 porciento de las células T mueren. Las que dejan el timo tienen restricción para el CPH. Las células T que reconocen autoantígenos son eliminadas o inactivadas.
Rearreglo de los genes del receptor de la célula T Los genes que codifican los RCT sufren rearreglos y las regiones V, de unión (J), de diversidad (D) y constante (C) se unen para constituir dos tipos de RCT, cada uno formado por dos cadenas. Cada tipo de RCT se asocia con una especificidad particular. Primero, los genes que codifican las cadenas g y d sufren rearreglos para formar el RCT-gd, después los genes que codifican las cadenas a y b se rearreglan para formar el RCT-ab. Después de ocurrida la combinación genética, el RCT se expresa en la superficie de la célula. Los RCT tienen aún más diversidad que las inmunoglobulinas porque tienen mayor número de genes para la región J y la región D, lo que permite codificar un repertorio preseleccionado de hasta 1015 especificidades. Al mismo tiempo que ocurre el rearreglo, se expresan varias moléculas accesorias importantes, incluyendo CD4, CD8, CD2 y CD34,5 [ver tabla 1]. (CD por las siglas en inglés de grupo de diferenciación). Relativamente pocas células T son CD4-, CD8- RCT-gd, cuya función no se conoce del todo. La mayoría se vuelven células RCT-ab que expresan en un principio CD4 y CD8 y que después, por selección positiva (ver adelante), se vuelven CD4+, CD8- RCT-ab (cooperadoras) o CD4-, CD8+ RCT-ab (que tienen funciones citotóxicas, supresoras o ambas). En todas las células T el CD3 se asocia con el RCT [ver figura 4].
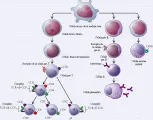
|
| Figura 4 |
| Maduración de los linfocitos T |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Selección positiva La selección positiva está controlada por células epiteliales de la corteza tímica.6 Debido a que las células T pueden reaccionar a los antígenos solo en asociación con el CPH propio, solo las células T con un RCT que puede fijarse al CPH propio se seleccionan. Cuando estas células reaccionan con el CPH propio de las células epitaliales del timo, las células de tipo CD4+, CD8+ RCT-ab que se fijan a moléculas del CPH de clase II se convierten en CD4+ y CD8-, reduciendo la expresión del CD8 y aumentando la del complejo RCT-CD3. Por el contrario, las células CD4+, CD8+ que se fijan a moléculas del CPH de clase I disminuyen la expresión de CD4 y se vuelven células CD4-, CD8+ RCT-ab. De este modo se seleccionan las células CD4+ RCT-ab y las células CD8+ RCT-ab con restricción para el CPH propio. Se supone que las células T que se unen, aún en forma débil, a las moléculas del CPH con ayuda del ligando CD4 o CD8, serán capaces de unirse con más intensidad a las células presentadoras de antígeno, por lo que no se requieren antígenos extraños para la selección positiva. El mecanismo de la selección positiva ha sido confirmado por estudios en los que se eliminó la molécula del CPH clase II en ratones7 y por experimentos en los que se emplearon ratones transgénicos para el RCT.8 La mayoría de las células T no interactúan con el CPH propio y sufren muerte programada, o apoptosis.
Selección negativa El proceso de selección negativa elimina las células T que tienen un RCT con afinidad intensa a los autoantígenos. Si no se eliminaran, estas células podrían causar enfermedades autoinmunes. Existen muchos autoantígenos en las células epiteliales del timo, y los estudios han demostrado que ciertos autoantígenos pueden ser presentados por varias células presentadoras de antígenos en el timo, como macrófagos y células dendríticas. Es posible que la selección negativa para algunos autoantígenos ocurra cuando las células T se trasladan hacia el sistema linfoide periférico después de dejar el timo. La interacción de alta afinidad entre el autoantígeno presentado por la molécula del CPH y el RCT en las células T inmaduras puede desencadenar la muerte celular. Por el contrario, la interacción entre un antígeno presentado por la molécula del CPH y el RCT de una célula T madura ocasiona activación celular.
Migración de las células T hacia los órganos linfoides secundarios Dos a tres días después de que las células tronco entran en el timo, los linfocitos migran a través de la pared de las venas poscapilares de la médula. Entran al torrente sanguíneo y se localizan en los órganos periféricos del sistema linfoide. Una vez ahí, los linfocitos dejan el torrente sanguíneo, de nuevo a través de las vénulas poscapilares, y entran a las regiones timo dependientes del sistema linfoide periférico, a la corteza interna de los ganglios linfáticos, a las vainas periarteriales del bazo y a las zonas intranodulares de las placas de Peyer, las amígdalas y el apéndice. Sin embargo, parece ser que la mayoría de las células T del intestino no derivan del timo, sino que se diferencían in situ en el epitelio intestinal. Estas células son CD4-, CD8- RCT-ab o células CD4-, CD8- RCT-gd.9
Si al momento de nacer el timo está ausente, el número total de linfocitos en sangre es menor que el normal y hay una carencia notable o ausencia de linfocitos T. Las áreas dependientes del timo del sistema linfoide periférico también carecen de linfocitos. Este estado se relaciona con una afección notable de las funciones inmunológicas mediadas por células, como la capacidad para resistir ciertas infecciones y para rechazar tejidos transplantados. Está aumentada la susceptibilidad para ciertos tumores; también existe depresión en las respuestas con anticuerpos que requieren de cooperación de los linfocitos T. Algunos de los efectos de la timectomía en animales de experimentación pueden ser parcialmente corregidos con extractos de timo. Se han aislado varias hormonas de probable estirpe tímica, incluyendo la timosina, la timulina y la timopoyectina. Se considera que estas hormonas favorecen la maduración de las células T e, in vitro, facilitan ciertas funciones de las células T.
El timo involuciona con la edad, lo cual podría explicar el desarrollo de deficiencias en el sistema inmunológico que ocurre en personas ancianas.
Tejido equivalente al de la bolsa y médula ósea
Además del timo, las aves cuentan con otro órgano linfoide primario, la bolsa de Fabricio, que también se deriva del epitelio intestinal en el embrión y está situada cerca de la cloaca. Las células tronco entran en la bolsa de Fabricio, donde se diferencían en linfocitos B, capaces de producir anticuerpos humonales. Por medio de manipulación quirúrgica u hormonal puede evitarse el desarrollo de la bolsa. Los pollos en quienes se ha efectuado extipación de la bolsa tienen una deficiencia completa de células B (y de células plasmáticas) y, por lo tanto, son incapaces de producir anticuerpos humorales. Sin embargo, estos animales son capaces de rechazar injertos y muestran reacciones mediadas por células; en comparación con las aves timectomizadas, muestran sólo una disminución mínima en el número de linfocitos en sangre. Este hallazgo indicaría que la mayoría de sus linfocitos circulantes son células T.
En los mamíferos no parece existir un órgano que sea equivalente a la bolsa de Fabricio. En la actualidad las evidencias sugieren que las células tronco pueden diferenciarse a células B en la médula ósea y en los órganos linfoides periféricos.
ORGANOS LINFOIDES SECUNDARIOS
Ganglios linfáticos
Los delicados capilares linfáticos situados en la periferia de los ganglios linfáticos pueden ser atravesados con facilidad por varias células errantes en el tejido conjuntivo, así como por macromoléculas exógenas y endógenas [ver figura 7]. Estas células y macromoléculas son transportadas a los ganglios linfáticos a través de vías aferentes. Los sinusoides de los ganglios linfáticos contienen macrófagos que pueden fagocitar antígenos y partículas. Los linfocitos circulan a través de los sinusoides y salen de los ganglios linfáticos por las vías eferentes. Por los linfáticos eferentes salen del ganglio un número 50 veces mayor de células, la mayor parte linfocitos, de los que ingresan por las vías aferentes. ¿De dónde provienen estas células? Menos del cinco porciento se deriva de precursores situados en los propios ganglios linfáticos. La mayor parte proviene de la circulación sanguínea, hecho que destaca la importancia de la recirculación linfocítica. La mayor parte de estos linfocitos sanguíneos son linfocitos T de vida prolongada que entran en los ganglios linfáticos a través de vénulas poscapilares situadas en la profundidad de la corteza. Estas células circulan por la corteza y siguen por la médula y los linfáticos eferentes. Unos cuantos linfocitos B también provienen de la circulación sanguínea; éstos parecen pasar en forma más indiscriminada a todas las áreas de los ganglios linfáticos, aunque algunos se localizan en los centros germinales de la corteza.
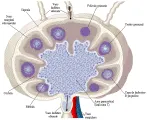
|
| Figura 7 |
| Estructura del ganglio linfático |
La mayor parte de los linfocitos B situados en los ganglios linfáticos se encuentran en los centros germinales de la corteza periférica. Cuando los ganglios linfáticos han sido estimulados por antígenos capaces de inducir la formación de anticuerpos, los centros germinales aumentan de tamaño y, por lo tanto, contienen muchos linfoblastos. A diferencia de las células T, la mayor parte de las células B no tienden a emigrar.
Los complejos antígeno-anticuerpo son captados por los macrófagos y por las células dendríticas foliculares en los senos subcapsulares de los ganglios linfáticos. Estas células dendríticas poseen muchos receptores para los componentes C3 del complemento y para la porción Fc de la IgG. Pueden retener el antígeno no degradado en la forma de complejos antígeno-anticuerpo-C3 durante meses. Es probable que este antígeno atrapado proporcione el estímulo para la formación de los centros germinales, donde proliferan las células B de memoria. Además, las células dendríticas foliculares producen cuerpos cubiertos de complejos inmunes denominados icosomas. Estos icosomas están dispersos en los centros germinales, se unen a las células B y sufren endocitosis.10 Entonces el antígeno es procesado y presentado a las células T. También se encuentran algunas células T en los centros germinales, en donde, a través de la producción de citocinas, ayudan a las células B estimuladas por un antígeno a proliferar y diferenciarse.
Las áreas paracorticales de los ganglios linfáticos contienen células interdigitantes (también denominadas dentríticas). Estas células tienen antígenos de clase II del CPH sobre su superficie y presentan el antígeno a las células T. Se considera que algunas de estas células interdigitantes se derivan de las células de Langerhans que adquieren el antígeno mientras están en la piel y posteriormente emigran a los ganglios linfáticos.
La formación de compartimientos en los ganglios linfáticos (centros germinales con linfocitos B inmóviles, áreas paracorticales con linfocitos T que viajan con rapidez, sinusoides llenos de macrófagos y la malla reticular de células dendríticas que tiende a mantener los antígenos por tiempo prolongado) parece facilitar las interacciones entre los diferentes tipos de células que se requieren para generar una respuesta inmunológica.
La función de los ganglios linfáticos puede estar afectada de forma importante en estados de inmunodeficiencia. Por ejemplo, cuando el timo está ausente existe una pérdida notable de linfocitos en las áreas paracorticales. En la agamaglobulinemia ligada al X hay pérdida completa de linfocitos B en los folículos germinales de los ganglios linfáticos y en ningún sitio se observan células plasmáticas.
Bazo
En el bazo los linfocitos se concentran en la pulpa blanca que rodea a las arteriolas centrales, formando las llamadas vainas linfoides periarteriolares [ver figura 8]. Los linfocitos T se encuentran sobre todo en la región central de estas vainas linfoides, mientras que los linfocitos B localizados en los centros germinales están más concentrados hacia la periferia. La pulpa blanca está separada de la pulpa roja por una zona marginal. Los linfocitos entran y salen de las vainas linfoides periarteriolares a través de los capilares de las arteriolas centrales en la zona marginal. Algunas células plasmáticas inmaduras pasan de la zona marginal a la pulpa roja.
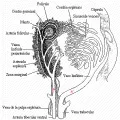
|
| Figura 8 |
| Estructura del bazo |
OTROS TEJIDOS LINFOIDES
Los linfocitos se encuentran también en otros lugares. El tejido linfoide relacionado con el intestino (TLRI) incluye las placas de Peyer y el apéndice. Estos tejidos contienen regiones con predominio de linfocitos T o B. En el intestino delgado también se encuentran muchos linfocitos, dentro de la lámina propia de la mucosa de las vellosidades y entre las células epiteliales de la mucosa. El tejido linfoide relacionado con las mucosas (TLRM) se encuentra en el aparato respiratorio, el intestino y el aparato urinario. Además, linfocitos aislados infiltran los tejidos epiteliales y conjuntivos de la mayor parte del organismo.
CIRCULACION DE LINFOCITOS
Existen tres tipos principales de circulación de los linfocitos: (1) la siembra de las células tronco provenientes del hígado fetal o de la médula ósea en los órganos linfoides primarios y la subsecuente diferenciación y distribución de estas células en el sistema linfoide periférico, (2) la recirculación sangre-linfa-sangre de linfocitos, y (3) la distribución de células efectoras a regiones particulares del organismo.
En el primer tipo de circulación, las células T tronco provenientes de la médula ósea penetran en el timo, donde proliferan y se diferencían en células inmunocompetentes en un plazo de dos a tres días; estas células son sembradas en los órganos linfoides periféricos. Además de las células T precursoras, la médula ósea también contiene algunas células T diferenciadas que son inmunocompetentes. La presencia de células inmunocompetentes en la médula ósea se demuestra en las notables reacciones de injerto contra huésped, mediadas por células T, que se presentan cuando se emplea médula ósea histoincompatible en la realización de trasplantes. Las células B tronco se movilizan de la médula ósea a los órganos linfoides secundarios. Los linfocitos B y T maduros no vuelven a entrar a los órganos linfoides primarios en cantidades significativas.
En el segundo patrón, la recirculación de linfocitos de la sangre, a través de los ganglios hacia los vasos linfáticos y de regreso a la sangre, es mucho más rápida y no entraña proliferación celular. Los tiempos promedio de tránsito de los linfocitos pequeños, la mayoría de los cuales son T con vida prolongada, son de 0.6 horas en la sangre, cinco a seis horas en el bazo, y 15 a 20 horas en los ganglios linfáticos. El tiempo de tránsito para los linfocitos B es más largo; por ejemplo, las células B pasan 30 horas en los ganglios linfáticos. Los linfocitos entran a los ganglios linfáticos a través de una zona especializada de la vénula poscapilar formada por células endoteliales altas y metabólicamente activas. Estas vénulas modificadas son denominadas vénulas endoteliales altas (VEA). Las citocinas producidas por las células T activadas cambian las vénulas planas por vénulas endoteliales altas. Los linfocitos se unen mejor a éstas últimas y después de unirse son capaces de viajar a través de las mismas.11
Las células T permanecen en la porción interna de la corteza, mientras que las B se mueven con lentitud hacia afuera, hasta los centros germinales. Se supone que en alguna parte de su trayecto las células B deben atravesar las áreas dependientes del timo y dirigirse hacia la médula, debido a que es ahí donde se encuentran la mayoría de las células plasmáticas (que se derivan de los linfocitos B). Si los linfocitos son extraídos del conducto tórácico sin permitir su reentrada a la sangre, las zonas dependientes del timo del sistema linfoide periférico se agotan de linfocitos mucho más rápidamente que los centros germinales; esta disminución selectiva es otra indicación de que las células B en estos órganos se mueven con más dificultad que las células T.
El tercer patrón de circulación de linfocitos es el movimiento de las células efectoras o de las células de memoria hacia los sitios de inflamación y hacia las mucosas. Las células T con marcador de superficie CD45RA se fijan en forma preferencial a las vélulas endoteliales altas de los ganglios linfáticos; por el contrario, las células T de memoria con el marcador de superficie CD45RO viajan de preferencia a la piel, las mucosas y otros tejidos normales, así como a sitios de inflamación. Los linfocitos tienen diversos receptores de superficie que influyen en el sitio al que se dirigen o donde se alojan. Estos receptores incluyen la L-selectina, el antígeno muy tardío-4 (VLA-4) y el antígeno asociado a la función del linfocito-1 (LFA-1). Las vénulas endoteliales altas de varios tejidos tienen también en su superficie moléculas de adhesión que contienen ligandos endoteliales selectivos para los tejidos, llamados adresinas vasculares. Estas adresinas, como la E-selectina, la molécula de adhesión intercelular-1 (ICAM-1), la ICAM-2, y la molécula de adhesión a células vasculares-1 (VCAM-1), se fijan a receptores específicos en los linfocitos. La expresión de estas adresinas es inducida por diversas citocinas.
Células inmunocompetentes de la respuesta inmunológica
Los linfocitos, las células principales en la respuesta inmunológica y las únicas células que reconocen antígenos específicos, constituyen una familia de células que tienen un núcleo redondo de localización central, carecen de gránulos específicos y tienen un citoplasma basófilo que contiene ribosomas libres. Las células T regulan la respuesta inmunológica, participan en las reacciones mediadas por células e inducen a los linfocitos B para producir anticuerpos. Las células B se diferencían en células plasmáticas productoras de anticuerpos. Ni los linfocitos T ni los B constituyen una población celular homogénea; cada grupo incluye diferentes subgrupos que pueden distinguirse por sus marcadores de superficie y por su función. Además, existe un grupo heterogéneo de linfocitos que no son ni células T ni B. Mientras que algunas de estas células pueden ser precursoras de las células T o B, otras parecen formar otro grupo o grupos generales. Estos grupos o subgrupos de linfocitos no pueden diferenciarse unos de otros por técnicas morfológicas. La fijación de anticuerpos monoclonales a marcadores de superficie es en la actualidad la técnica más específica para distinguir entre los subgrupos de linfocitos.
CELULAS T
La observación de que algunos linfocitos humanos pueden fijar eritrocitos de carnero, formando rosetas, llevó a la designación del marcador original de los linfocitos T humanos. El uso de anticuerpos monoclonales reveló que los eritrocitos de carnero se fijan en una glucoproteína de 55 kd situada sobre la superficie del linfocito, en la actualidad este antígeno se denomina CD2. Todas las células T maduras y los timocitos medulares expresan el antígeno CD2, así como otro antígeno de superficie, conocido como CD3. Los linfocitos portadores de CD2 están ausentes o muy disminuidos en la sangre de los niños con aplasia tímica, pero aparecen con rapidez después de colocar un injerto de timo.
Subgrupos de células T
Se han identificado varios subgrupos de células T que tienen diferentes funciones y se les ha analizado empleando una serie de anticuerpos monoclonales dirigidos contra las moléculas de superficie.12 Del 50 al 65 porciento de las células T periféricas muestran una proteína de superficie de 57 kd conocida como CD4, la cual se reconoce por los anticuerpos monoclonales OKT4 y anti-Leu-3. Del 25 al 35 porciento de las células T periféricas muestran una glucoproteína de 32 kd llamada CD8, que se reconoce por los anticuerpos monoclonales OKT8 y anti-Leu-2. Aunque CD4 y CD8 se expresan juntas en los timocitos en la etapa II, sólo una u otra se expresa en los subgrupos complementarios de las células T maduras (células T CD4+ y CD8+). Las células T CD4+ reconocen antígenos cuando se presentan asociados con las moléculas de clase II del CPH (antígenos HLA-D); las células T CD8+ reconocen antígenos en la vecindad de moléculas de clase I del CPH (antígenos HLA-A, HLA-B y HLA-C).
Las moléculas CD4 y CD8 se acoplan a una tirosina cinasa específica denominada p56lck en el citoplasma de la célula T.12 Esta cinasa se relaciona con la familia del protooncogen src que está implicado en la transformación y activación de algunas células de mamíferos, y cinasas similares forman parte de la cola citoplasmica de receptores para hormonas de crecimiento, como el factor de crecimiento epidémico, el factor de crecimiento derivado de las plaquetas y la insulina. En estudios in vitro, los complejos CD4-p56lck y CD8-p56lck fosforilaron la porción citoplásmica de las subunidades g, d, e, y en especial la subunidad s de CD3, pero no fosforilaron otros 12 antígenos CD probados.12 Esta interacción explica cómo CD4 y CD8 interactúan con el complejo CD3-RCT y pueden desempeñar un papel importante en la activación de la célula T. El papel crucial de la derivación de CD4-RCT en la señal para la activación de la célula T se apoya en otros dos estudios. Es interesante que el antígeno común leucocitario CD45 parece tener un dominio intracelular con actividad de fosfatasa de tirosina proteica y podría regular la activación de las células T por la cinasa p56lck.
Las células T CD4+, pueden diferenciarse en células TH1 y TH2 según las linfocinas que producen.13,14 Las células TH1 producen interleucina-2 (IL-2) e interferón gama, que son importantes en las reacciones inmunológicas mediadas por células. Las células Th2, que producen interleucina-4 (IL-4), interleucina-5 (IL-5), interleucina-6 (IL-6) e interleucina-10 (IL-10), ayudan a las células B en la producción de anticuerpos. Las linfocinas que son producidas por cada uno de estos tipos celulares influyen en el otro tipo celular. El interferón gama producido por las células TH1 puede inhibir la función de las células TH2, mientras que la IL-10, que es producida por células TH2 (así como por monocitos, macrófagos y células B), puede inhibir la función de las células TH1. La IL-10 no actúa en forma directa sobre los TH1 sino sobre células accesorias de las que éstos dependen. Las citocinas formadas por otras células también pueden influir en las subpoblaciones de linfocitos T. Por ejemplo, la interleucina-12 (IL-12) favorece la diferenciación de las células TH1.
CELULAS B
Los linfocitos B son las células productoras de inmunoglobulinas del sistema inmune, y pueden ser identificadas por la presencia de inmunoglobulinas en su superficie. Estas células con receptores para inmunoglobulinas positivos en la superficie de su membrana (SmIg+) y constituyen del 5 al 15 porciento de los linfocitos en sangre periférica [ver figura 11]. Esta definición no incluye a los precursores de las células B, que tienen IgM en su citoplasma pero no en su superficie, ni a las células plasmáticas SmIg- que se originan a partir de las células B. La mayor parte de las células B tienen IgM e IgD en su superficie y cerca de la cuarta parte del total de estas células sólo poseen inmunoglobulinas de alguna de estas dos clases. Uno porciento de las células B muestra IgG o IgA Los linfocitos T y B constituyen del 80 al 95 porciento de los linfocitos en sangre periférica. El resto de las los linfocitos presentes en sangre periférica constituyen una población heterogénea de células (ver adelante).
|
| Figura 11 |
| Patrón de inmunofluorescencia de inmunoglobulinas en el linfocito |
Sobre la superficie de las células B se encuentra un receptor del complemento llamado receptor 2 del complemento (RC2), el cual se une a los factores C3d, C3dg y al factor iC3b. Las células B también poseen receptores para el virus de Epstein-Barr y para la porción Fc de la IgG. La molécula CD20, una fosfoproteína de 35 kd que se encuentra sobre la superficie de las células B, es un marcador de los linfocitos B utilizado con frecuencia. Algunos aloantígenos están presentes sobre la superficie de los linfocitos B, pero no sobre la superficie de los linfocitos T. Estos antígenos se detectan usando sueros de mujeres embarazadas y algunos antisueros para antígenos purificados obtenidos de las líneas celulares de linfoblastos B. Algunos precursores de las células B carecen del receptor SmIg, pero poseen estos aloantígenos.
Un grupo de células B, las células de memoria, aumentan la respuesta inmune a los antígenos ya conocidos, y constituyen la llamada respuesta inmune secundaria, respuesta anamnésica o respuesta de refuerzo. Estas células B de memoria sufren mutaciones somáticas en las regiones variables de sus genes de inmunoglobulina. Cuando esta mutación somática ocurre en los centros germinales de los ganglios linfáticos que contienen antígenos unidos a las células dendríticas foliculares, se presenta selección de las células B de memoria que tienen receptores de alta afinidad para los antígenos.
Un subgrupo de células B, los llamados linfocitos B1, se desarrollan en fases tempranas y tienen vida media muy prolongada. Los experimentos realizados sobre la capacidad de los progenitores de células B para reconstituir diferentes poblaciones de linfocitos B en ratones adultos radiados sugieren que las células B1 y los linfocitos B regulares se producen a través de linajes diferentes.15 Se encuentran progenitores de células B1 en el higado fetal y en el epiplón embrionario, pero no en la médula ósea adulta. Las células B1 que expresan CD5 en su superficie se conocen como B1a, y las que no se denominan células B1b. Las células B1 se asocian con frecuencia con producción de autoanticuerpos. También elaboran grandes cantidades de IL-10.
Al hacer el recuento de las células SmIg+ es importante recordar que las inmunoglobulinas producidas por las células B pueden fijarse de manera pasiva a la superficie de las membranas de células no-B. Por ejemplo, las células que tienen receptores Fc fijan complejos de inmunoglobulina; más aún, los anticuerpos fluorescentes para inmunoglobulinas, que se emplean con frecuencia para identificar a las células SmIg+, también pueden fijarse a los receptores Fc. Este problema puede evitarse empleando anticuerpos fluorescentes F(ab')2, esto es, la inmunoglobulina cuya fracción Fc ha sido eliminada. Además, la presencia de autoanticuerpos contra linfocitos T incrementará el número de células SmIg+ que son confundidas con células B. Estos anticuerpos antilinfocíticos se han encontrado en varias enfermedades, incluyendo el lupus eritematoso generalizado (LEG), la mononucleosis infecciosa, la neumonía atípica, la artritis reumatoide y varios trastornos linfoproliferativos. La presencia de estos anticuerpos se sospecha por lo general cuando los porcentajes de células CD3+ y células SmIg+ suman más del 100 porciento. Las inmunoglobulinas absorbidas de manera pasiva pueden eliminarse incubando las células toda una noche en ausencia de suero humano o por medio de un ligero tratamiento con tripsina. En este último caso las células se cultivan poco tiempo después del tratamiento con tripsina para que las células B genuinas puedan regenerar su SmIg.
CELULAS ASESINAS NATURALES
Las células asesinas naturales (NK, por sus siglas en inglés, n. del t.) son linfocitos granulares de gran tamaño que no poseen el complejo RCT-CD3 característico de las células T ni el SmIg de las células B. In vitro, estas células pueden destruir diversas células tumorales o infectadas por virus de modo no específico. Esto es, no requieren de una sensibilización previa o de la presencia de anticuerpos para ser citotóxicas. Sus gránulos contienen proteínas formadoras de poros que pueden mediar la lisis celular. Las células NK no tienen memoria ni muestran restricción de CPH. No se conoce con precisión al precursor de las células NK, pero se piensa que es diferente a la célula tronco tímica (precursora de los linfocitos T) y a la célula pro-B (precursora de los linfocitos B).
Las células asesinas naturales poseen varios marcadores de superficie, incluyendo el CD56, una glucoproteína de 140 kd también denominada NKH1, el CD2, que también está presente en las células T y el CD16, un receptor Fc. El CD16 se asocia con la cadena s del complejo RCT-CD3. Esta cadena participa en la transducción de señales desencadenada por los anticuerpos que se fijan al CD16, lo que activa la célula.
La IL-12 estimula la proliferación de las células NK y su producción de interferón gama, que es importante en muchas reacciones inmunológicas.16
Una población de células T citotóxicas no restringida al CPH puede lisar algunas de las mismas células blanco que destruyen las células asesinas naturales. Estas células T citotóxicas tienen un marcador de superficie que puede detectarse por medio del anti-CD56, pero no por anti-CD16. Además, expresan el complejo RCT-CD3.17
Una pequeña proporción de las células nulas de la sangre son precursores mieloides y precursores de los linfocitos T y B inmaduros.
CELULAS PLASMATICAS
Por la influencia de los antígenos, los linfocitos T y células accesorias, las células B se diferencian en células plasmáticas, que son células maduras productoras de anticuerpos. Estas células son de mayor tamaño que los linfocitos y se caracterizan por un núcleo redondo y excéntrico con heterocromatina gruesa distribuida en un patrón en rueda de carreta. Las células plasmáticas tienen un citoplasma altamente basófilo, y un retículo endoplásmico bien desarrollado, con frecuencia organizado en capas concéntricas. Las células plasmáticas pueden estar repletas de material granular, que corresponde al anticuerpo que están produciendo [ver figura 12]. Algunas veces, una o más de las cisternas del retículo endoplásmico se encuentran distendidas por grandes inclusiones llamadas cuerpos de Russell; éstos son agregados de moléculas de inmunoglobulinas formadas de manera incompleta. Las células plasmáticas no tienen inmunoglobulinas en su superficie. Estas células son terminales, lo que significa que no se dividen. Los precursores inmaduros de las células plasmáticas, los plasmoblastos, son difíciles de distinguir de los linfoblastos y de los linfocitos grandes. En condiciones normales no se encuentran células plasmáticas en sangre periférica.
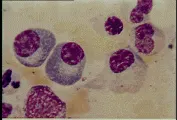
|
| Figura 12 |
| Células plasmáticas |
MONOCITOS Y MACROFAGOS
Los monocitos pertenecen al sistema fagocítico mononuclear, antes llamado sistema reticuloendotelial. Son grandes células mononucleares que constituyen del tres al ocho porciento de los leucocitos en sangre periférica. Su citoplasma es mucho más abundante que el de los linfocitos. Su núcleo es a menudo excéntrico y oval o reniforme [ver figura 13]. Los lisosomas llenos de enzimas degradantes aparecen como pequeñas vacuolas en el citoplasma. Los monocitos se originan a partir de los promonocitos, los cuales son precursores con una tasa elevada de división y que se encuentran en la médula ósea. Cuando las células maduras entran a la sangre periférica son llamadas monocitos; cuando abandonan la sangre e infiltran los tejidos, sufren cambios adicionales y entonces son conocidas como macrófagos.
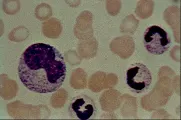
|
| Figura 13 |
| Monocito |
Los macrófagos desempeñan un papel importante en la inducción de la respuesta inmunológica al ser uno de los tipos de células que presenta el antígeno a los linfocitos. También actúan como células efectoras, atacando ciertos microrganismos y células neoplásicas, y eliminando material extraño.
Los macrófagos contienen receptores específicos para anticuerpos y complemento, lo que incrementa su capacidad para fagocitar microrganismos cubiertos con estas moléculas. Los receptores de anticuerpos son para las porciones Fc de la IgG1 y de la IgG3; existe también un receptor Fc para la IgE. Tienen dos receptores del complemento: el receptor 1 (RC1) y el receptor 3 (RC3). El RC1 tiene alta afinidad por el componente C3b del complemento y menor afinidad por iC3b y C4b. El RC3, también llamado MAC-1, interactúa con iC3b y con ciertas moléculas de carbohidratos, incluyendo los antígenos que contienen carbohidratos del protozoario Leishmania.
Los macrófagos poseen también receptores para citocinas; el interferón gama es la principal citocina activadora de estas células.
Una pequeña proporción de los antígenos de clase II del CPH está presente en los monocitos; la expresión de las moléculas de clase II del CPH se incrementa considerablemente cuando se activan los macrófagos, lo cual puede ocurrir por varias linfocinas, incluyendo el interferón gama, el factor estimulador de colonias de granulocitos y macrófagos (FEC-GM), el factor activador de los macrófagos (FAM) y el factor inhibidor de la migración (FIM). Linfocinas como la interleucina-4 y el factor-b transformador del crecimiento (FCT-b) antagonizan esta activación. El antígeno Mo3e, una proteína de 50 kd, es un receptor de urocinasa. Este antígeno ha sido identificado por medio de anticuerpos monoclonales y parece ser específico de los macrófagos activados. El Mo3e no se ha encontrado en otras células hematopoyéticas.18
Los macrófagos producen un gran número de sustancias solubles que son importantes en la respuesta inmunológica y en el proceso de inflamación.19 Estas sustancias incluyen enzimas como el factor activador del plasminógeno y la elastasa, factores de crecimiento como el FEC-GM, citocinas como la interleucina-1 (IL-1), IL-6, IL-10, IL-12 y el factor de necrosis tumoral-a (FNT-a), factores que son críticos para combatir a los microrganismos, como los metabolitos del oxígeno y el óxido nítrico, componentes del complemento de la vía clásica y alterna, proteínas inflamatorias de macrófagos (PIM), y factores que facilitan la reparación tisular, como el factor de crecimiento de fibroblastos (FCF).
GRANULOCITOS
Neutrófilos
El granulocito predominante en la sangre es el neutrófilo. Se origina a partir de precursores en la médula ósea, su vida media en sangre es de cuatro a ocho horas, pero en los tejidos vive hasta tres veces más. Los neutrófilos son atraídos de la sangre hacia los lugares en donde hay inflamación por diversos factores quimiotácticos, incluyendo aquellos generados por activación de la cascada del complemento y algunos producidos por linfocitos estimulados por antígenos. Los neutrófilos tienen una importancia primaria en la defensa del organismo debido a su capacidad para fagocitar y destruir microrganismos. Este proceso es facilitado considerablemente por el complemento y los anticuerpos. La importancia de la fagocitosis por neutrófilos se ilustra de manera especial por el hecho de que en pacientes con deficiencia o ausencia total de estas células las infecciones piógenas ponen en peligro la vida.
Los neutrófilos también contienen gránulos lisosomales llenos de una variedad de enzimas degradantes y de pequeñas proteínas que pueden causar inflamación cuando son liberadas por las células. Muchos neutrófilos se encuentran en los lugares donde se localizan ciertas reacciones inmunopatológicas, en especial aquellas desencadenadas por complejos inmunes. Estas células están dotadas con receptores para IgG y para el componente C3b del complemento.
Eosinófilos
Los eosinófilos se caracterizan por poseer grandes gránulos rojos, que contienen proteínas básicas y muchas enzimas degradantes. La función exacta de los eosinófilos ha sido desde hace mucho tiempo un misterio, aunque se han sugerido varias funciones diferentes. Por ejemplo, se han encontrado muchos eosinófilos en tejidos donde se presentan reacciones inmunológicas que han sido desencadenadas por anticuerpos IgE, como en los pólipos nasales o en la pared bronquiolar de algunos enfermos con asma. Los eosinófilos son activados cuando el antígeno estimula a las células cebadas cubiertas con IgE para liberar un factor quimiotáctico de la anafilaxia para eosinófilos. El hallazgo de que los eosinófilos contienen diversas enzimas que pueden degradar a los mediadores de la hipersensibilidad inmediata, como la histamina, la sustancia de rección lenta de la anafilaxia (leucotrienos) y el factor agregante plaquetario, sugiere que dichas células pueden controlar o disminuir tales reacciones de hipersensibilidad. Los eosinófilos también son atraídos por los complejos antígeno-anticuerpo y pueden fagocitarlos. Algunos eosinófilos tienen receptores para Fc y C3b lo que les permite unirse a los complejos antígeno-anticuerpo o a los organismos que han desencadenado la vía alterna del complemento y están cubiertos con C3b.
Los eosinófilos se acumulan con frecuencia en sitios de infecciones producidas por helmintos. En la esquistosomiasis, por ejemplo, los eosinófilos se encuentran alrededor de las larvas y en los granulomas inducidos por los huevos. Debido a que los eosinófilos modulan la citotoxicidad mediada por células dependientes de anticuerpos (CCDA), se considera que participan en la resistencia a algunas infecciones parasitarias. Los eosinófilos pueden también contribuir a la inflamación y a cambios tisulares como los observados en pacientes con fibrosis endomiocárdica que se desarrolla después de hipereosinofilia prolongada. La capacidad de los eosinófilos para destruir larvas cubiertas con anticuerpos está aumentada por el factor reforzador citotóxico eosinofílico derivado de los monocitos, así como por el FNT-a y el FEC-GM. La liberación de estas citocinas por los macrófagos, células cebadas o linfocitos probablemente aumenta la inflamación producida por los eosinófilos. La IL-5 es crucial en la produción de eosinofilia; en los ratones, los anticuerpos anti-IL-5 disminuyen notablemente el número de eosinófilos en la sangre y los tejidos.
Basófilos y células cebadas
Los basófilos y las células cebadas tienen gránulos citoplásmicos metacromáticos. La estructura de los gránulos está formada por heparina unida a diversas aminas vasoactivas y otros mediadores. Los basófilos tienen un núcleo multilobulado y relativamente pocos gránulos metacromáticos, de gran tamaño, mientras que las células cebadas tienen un núcleo unilobular y gránulos citoplásmicos pequeños y más numerosos. Los basófilos se encuentran sobre todo en la sangre y en sitios de inflamación, mientras que las células cebadas están presentes principalmente en los tejidos. Aunque las dos células tienen funciones en común, se originan de diferentes precursores. Las células cebadas tienen más enzimas en sus gránulos que los basófilos y pueden liberar varias citocinas, incluyendo interleucina-3 (IL-3), IL-4, IL-5, IL-6 y FEC-GM. Ambos tipos de células tienen receptores Fc para IgE e IgG; de hecho, es la combinación del antígeno con la IgE ligada a estas células la responsable de la liberación de los mediadores en las reaciones de hipersensibilidad inmediata. Se ha demostrado que las células cebadas también poseen receptores para C3b.
Además de su papel en las reacciones de hipersensibilidad inmediata, los basófilos se encuentran en gran número en algunas reacciones tardías mediadas por células, como la hipersensibilidad por contacto en humanos, el rechazo a tumores o a injertos de piel y la hipersensibilidad a ciertos microrganismos en animales.
PLAQUETAS
La principal función de las plaquetas es participar en la coagulación sanguínea, pero también están implicadas en algunos de los mecanismos secundarios iniciados por la respuesta inmune. Las plaquetas poseen receptores Fc para la IgG e IgE y receptores para las moléculas de clase I del CPH sobre su superficie. Se adhieren y se agregan sobre las superficies endoteliales dañadas, liberando factores que producen aumento de la permeabilidad capilar y activan diversos componentes del complemento que en forma secundaria atraen leucocitos al lugar de la lesión. Bajo ciertas circunstancias se ha demostrado que las plaquetas participan en la defensa del huésped contra microrganismos invasores.
- Ehrlich P, Morgenroth J: Ueber hämolysine. Klin Wochenschr 38:251,1901
- Bumet M: The Clonal Selection Theory of Acquired Immunity. Cambridge University Press, London, 1959
- Miller JF AP , Osoba D: Current concepts of the immunological function of the thymus. Physiol Rev 47:437, 1967
- Knapp W, Dorken B, Gilks WR, et al, Eds: Leukocyte Typing IV: White Cell Differentiation Antigens. Oxford University Press, Oxford, England, 1989
- SchIossman SF, Boumsell L, Gilks W, et al: Update: CD antigens 1993. J ImmunoI 152:1, 1994
- Cosgrove D, Chan SH, Waltzinger C,etal: The thymic compartment responsible for positive selection of CD4 + T cells. Int lmmunol4:707 , 1992
- Grusby MJ, Johnson RS, Papaioannou VE, et al: Depletiofl of CD4+ T cells in major histocompatibility complex class II-deficient mice. Science 253:1417,1991
- Sha WC, Nelson CA, Newberry RD, et al: Positive and negative selection of an antigen receptor on T cells in transgenic mice. Nature 336:73, 1988
- Poussier P, Edouard P, Lee C,et al: Thymus-independent development and negative selection of T cells expressing T cell receptor a/in the intestinal epithelium: evidence for distinct circulation patterns of gut- and thymus-derived T lymphocytes. J Exp Med 176:187,1992
- Tew JG, Koscoe MH, szakal AK: The altemative antigen pathway. Immunology Today 10:229,1989
- Picker LJ, Butcher EC: Physiological and molecular mechanisms of lymphocyte homing. Annu Rev Immunol 10:561,1992
- Rudd CE, Anderson P, Morimoto C, et al: Molecular interactions with T subsets and a role of the CD4/CD8: p56lck complex in human T cell activation. Immunology Reviews 111:255, 1989
- Mosmann TR, Coffman RL: Heterogeneity of cytokine secretion patterns and functions of helper T cells. Adv Immunol 46:111, 1989
- Romagnani S: Human THl and TH2 subsets: doubt no more. Immunol Today 12:256, 1991
- Kantor AB, Herzenberg LA: Origin of murine B cell lineages. Annu Rev Immunoll 1:501, 1993
- WoIf SF, Temple PA, Kobayashi M, et al: Cloning of cDNA for natural killer cell stimulatory factor, a heterodimeric cytokine with multiple biologic effects on T and natural killer cells. J Immunol 146:3074,1991
- Lanier LL, Le AM, Cwirla S, et al: Antigenic, functional, and molecular genetic studies of human natural killer cells and cytotoxic T lymphocytes not restricted by the major histocompatibility complex. Fed Proc 45:2823,1986
- Todd RF III, Liu DY: Mononuclear phagocyte activation: activationassociated antigens. Fed Proc 45:2829,1986
- Auger MJ, Ross JA: The bilogy of the macrophage. The Macrophage. Lewis CE, McGee JO'D, Eds. Oxford University Press, Oxford, England, 1992, p 1