Hematología
⭳ Abrir artículo (PDF)813.5 KBEste artículo fue revisado respecto a la Edición 3/2000. Ver esa versión →
Contenido del artículo
II FUNCION DEL ERITROCITO Y TRASTORNOS DEL METABOLISMO DEL HIERRO
- Función del eritrocito
- ESTRUCTURA DE LA HEMOGLOBINA
- CURVA DE DISOCIACION DE OXIGENO-HEMOGLOBINA
- EFECTOS DEL 2,3-DIFOSFOGLICERATO
- APORTE DE OXIGENO A LOS TEJIDOS
- TRANSPORTE DE BIOXIDO DE CARBONO
- Trastornos del metabolismo del hierro
- METABOLISMO DEL HIERRO
- BALANCE DEL HIERRO
- TRANSPORTE, ALMACEN Y REGULACION DEL HIERRO
- TRANSFERRINA
- RECEPTOR DE TRANSFERRINA
- FERRITINA
- PROTEINAS REGULADORAS DE HIERRO
- APORTE Y ALMACEN DE HIERRO EN LA CELULA
- REGULACION DE LA CAPTACION DE HIERRO EN LA CELULA Y SU ALMACEN POR PROTEINAS REGULADORAS DE HIERRO
- Evaluación de laboratorio del metabolismo del hierro
- Exceso de hierro
- EXCESO DE HIERRO CAUSADO POR MAYOR ABSORCION
- Hemocromatosis hereditaria (asociada al HLA)
- Anemias con exceso de hierro
- Otras formas de exceso de hierro causadas por aumento en la absorción
- EXCESO DE HIERRO POR TRANSFUSIONES Y ADMINISTRACION PARENTERAL
- OTRAS FORMAS DE EXCESO DE HIERRO
- Deficiencia de hierro
II FUNCION DEL ERITROCITO Y TRASTORNOS DEL METABOLISMO DEL HIERRO
DR. GARY M. BRITTENHAM
Función del eritrocito
La función principal del eritrocito es el transporte de oxígeno de los pulmones a los tejidos periféricos para su utilización y del bióxido de carbono a los pulmones para su excreción.1,2 El eritrocito maduro dedica más del 95 porciento de su proteína intracelular, la hemoglobina, a esta tarea. La hemoglobina, que es la molécula transportadora del oxígeno, se une a las moléculas de oxígeno en el ambiente de alta tensión de oxígeno de los alveolos pulmonares y libera moléculas de oxígeno en los tejidos periféricos en donde la tensión de oxígeno es baja. En el eritrocito, se encuentra dentro de una bicapa lípida sostenida por un citoesqueleto que determina la forma bicóncava y discoide de esta célula. La forma del eritrocito optimiza el paso de la célula a través del sistema circulatorio y permite la aposición de los eritrocitos y las células parenquimatosas a través del delgado endotelio de los capilares, lo que facilita el intercambio de oxígeno y bióxido de carbono. Sin eritrocitos, la sangre del plasma transportaría solo alrededor de 5 ml de O2/L, pero con una hemoglobina normal la sangre total puede transportar alrededor de 200 ml de O2/L.2
ESTRUCTURA DE LA HEMOGLOBINA
La hemoglobina, que tiene un peso molecular de 64,400 daltons, es un tetrámero compuesto de dos pares de cadenas polipeptídicas globinas diferentes con un grupo hem, la ferroprotoporfirina IX, unido en forma covalente a un sitio específico en cada cadena.3 La principal hemoglobina adulta, la hemoglobina A, está formada por un par de cadenas a (cada una conteniendo 141 aminoácidos) y un par de cadenas b (cada una con 146 aminoácidos), y se expresa como a2b2. Su estructura tridimensional es esferoidal, con un solo eje de asimetría y los cuatro grupos hem en surcos colocados equidistantes alrededor de la superficie de la molécula.4 La configuración deoxi de la hemoglobina se estabiliza por uniones intra e intercadenas, algunas de estas dependen de la unión de protones y otras de la presencia de 2,3-difosfoglicerato (2,3-DPG). Estas uniones se rompen en forma secuencial durante la oxigenación. Con la unión de una molécula de oxígeno a un grupo hem cada subunidad a o b no unida sufre un cambio en su estructura terciaria, lo que produce un cambio en la configuración de toda la molécula de hemoglobina [ver figura 1]. Estos cambios de conformación producen un aumento en la afinidad por oxígeno del resto de las subunidades no unidas, un fenómeno denominado cooperatividad o interacción hem-hem.5,6 Por el contrario, si el oxígeno es liberado de un hem en la oxihemoglobina, el resto de los sitios unidos sufren cambios que disminuyen su afinidad por el oxígeno, facilitando la liberación de esta molécula. Estos cambios conformacionales son también los responsables de la menor afinidad de la hemoglobina por el oxígeno al disminuir el pH, el llamado efecto Bohr.3 La oxigenación disminuye los valores K de los grupos ácidos en residuos específicos de aminoácidos, liberando alrededor de 2.8 protones por molécula de hemoglobina. El efecto Bohr es fisiológicamente benéfico tanto en los pulmones, en donde la liberación de bióxido de carbono aumenta el pH, aumentando la afinidad por oxígeno y su captación, como en los tejidos, en donde la captación del bióxido de carbono disminuye el pH, reduciendo la afinidad por oxígeno y facilitando su liberación.
CURVA DE DISOCIACION DE OXIGENO-HEMOGLOBINA
La curva de disociación de oxígeno-hemoglobina [ver figura 2] es una gráfica del equilibrio entre el oxígeno y la hemoglobina a varias tensiones de oxígeno (PO2). La forma sigmoidea de la curva, que es resultado de la interacción entre las subunidades de la hemoglobina humana, tiene ventajas fisiológicas.1-3 En los pulmones, con una presión parcial de oxígeno de alrededor de 90 mm Hg, la hemoglobina está saturada al 97 porciento. Después de dejar el oxígeno en los tejidos, a una PO2 de alrededor de 40 mm Hg en la sangre venosa mixta, la saturación de la hemoglobina es de 75 porciento. Por lo tanto, se libera una cantidad relativamente grande de oxígeno con una disminución discreta en la tensión de oxígeno. La P50, que es la presión parcial de oxígeno a la que la hemoglobina está saturada a la mitad, es una medida útil de la afinidad de la hemoglobina por el oxígeno: a mayor afinidad menor P50. En condiciones fisiológicas (i.e., temperatura de 37C [98.6F], pH de 7.40, 2,3-DPG de 5 mmol/L y tensión de bióxido de carbono [PCO2] de 40 mm Hg), la P50 de la sangre del adulto normal es de 26±1 mm Hg. La P50 disminuye (se desplaza a la izquierda) con el aumento en el pH, la reducción en el 2,3-DPG o la reducción en la temperatura.
EFECTOS DEL 2,3-DIFOSFOGLICERATO
El producto intermedio de la glucólisis, 2,3-DPG, un anión con carga importante, está presente en los eritrocitos maduros en una concentración de 4 a 5 mmol/L, aproximadamente la misma concentración celular que la hemoglobina. Una molécula de 2,3-DPG se une a una molécula de desoxihemoglobina en el hueco formado por los extremos amino de las cadenas b, reduciendo la afinidad del oxígeno por la hemoglobina al estabilizar la conformación desoxi1-3 sin alterar la interacción o el efecto Bohr. El trifosfato de adenosina (ATP) se une a la hemoglobina en forma semejante, pero la mayoría del ATP intracelular forma un complejo con Mg2+, que bloquea la reacción con la desoxihemoglobina. Como resultado, el 2,3-DPG es el regulador alostérico cuantitativamente más importante de la afinidad por el oxígeno.2 En presencia de hipoxia (por anemia aguda, enfermedad cardiopulmonar, otros trastornos o exposición súbita a altitudes elevadas), la concentración de 2,3-DPG aumenta en horas, incrementando la P50. El desplazamiento de la curva de disociación de oxígeno-hemoglobina a la derecha facilita la entrega de oxígeno a los tejidos, pero a costa de disminuir su adquisición en los pulmones. Los aumentos en el 2,3-DPG pueden ayudar a adaptarse a corto plazo al estrés hipóxico si el aporte de oxígeno es abundante y la reserva cardiopulmonar robusta. A altitudes elevadas, cuando el sistema cardiovascular es incapaz de satisfacer en forma eficaz la mayor demanda circulatoria, o en otras circunstancias patológicas, el aumento en la cantidad de 2,3-DPG puede ser contraproducente.
APORTE DE OXIGENO A LOS TEJIDOS
El eritrocito circulante es un elemento crucial en el transporte de oxígeno a los tejidos, pero una variedad de otros componentes fisiológicos deben funcionar también en forma integrada para proporcionar un aporte adecuado de oxígeno.2 El volumen y la viscosidad de la sangre afectan el aporte de oxígeno. La concentración de eritrocitos circulantes depende, en parte, de la actividad eritropoyetica, que a su vez es regulada por la producción renal de eritropoyetina. Otros componentes importantes incluyen la tensión de oxígeno en el ambiente, los factores pulmonares que influyen en la oxigenación y la ventilación, factores cardiovasculares como el gasto cardiaco y el flujo sanguíneo regional y la magnitud de oxígeno que no se libera en los tejidos.
TRANSPORTE DE BIOXIDO DE CARBONO
La hemoglobina se une al bióxido de carbono después de dejar oxígeno en los tejidos. La mayoría del bióxido de carbono de los capilares tisulares es transportado a los pulmones como bicarbonato, y alrededor de un 10 porciento es transportado como un complejo carbamino unido en forma reversible a los grupos aminoterminales de las cadenas de globina. La desoxihemoglobina tiene mayor afinidad por el bióxido de carbono que la oxihemoglobina, lo que facilita su transporte desde los tejidos para eliminarse en el pulmón.
Trastornos del metabolismo del hierro
Todas las células del organismo requieren hierro. En el metabolismo humano el hierro se usa para transportar y almacenar oxígeno, transportar electrones y actuar como el elemento catalítico cuantitativamente más importante, con funciones cruciales en el metabolismo oxidativo y en el crecimiento y proliferación celular. En los humanos el hierro se encuentra casi en forma exclusiva como ión ferroso (Fe2+) o férrico (Fe3+) y puede cambiar con facilidad entre estos dos estados de oxidación. Debido a que estas formas del hierro son muy reactivas, casi siempre existe como un componente de complejos hierro-porfirina o metaloenzimas, o unido a proteínas especializadas de transporte o almacén. Estos compuestos evitan o limitan la aparición de hierro en forma no unida o inorgánica, evitando la participación del hierro en reacciones de producción de radicales libres que pueden lesionar los componentes celulares. Bajo condiciones fisiológicas, la cantidad de hierro en el organismo está controlada dentro de límites estrechos para evitar o su deficiencia o su exceso. Cuando existe deficiencia de hierro el organismo es incapaz de sintetizar suficiente cantidad de hem, otros complejos hierro-porfirina, metaloenzimas u otros compuestos que contienen hierro para realizar las funciones normales. Con la sobrecarga de hierro, el exceso puede provocar reacciones dañinas que causan daño progresivo y eventualmente letal a órganos vitales.
METABOLISMO DEL HIERRO
Normalmente la concentración de hierro en el organismo es de 40 mg Fe/kg de peso corporal en la mujer y alrededor de 50 mg Fe/kg en el hombre. La mayoría de este hierro es un componente de varios compuestos funcionales dedicados al transporte, entrega, utilización o consumo de oxígeno. La mayor proporción de hierro se encuentra en el eritrón como hierro de la hemoglobina (28 mg/kg en mujeres y 32 mg/kg en hombres), dedicado al transporte y entrega de oxígeno. Se encuentran también cantidades pequeñas de hierro eritrón (<1mg/kg) en el hem y en enzimas no hem en eritrocitos en desarrollo.
El resto del hierro funcional se encuentra como hierro de mioglobina (4 mg/kg en mujeres y 5 mg/kg en hombres) en el músculo y en enzimas que contienen o dependen de hierro (1 a 2 mg/kg) en todas las células del organismo. Se depositan pequeñas cantidades de hierro dentro de ferritina en las células eritroides, pero el principal almacen de hierro (5 a 6 mg/kg en mujeres y 10 a 12 mg/kg en hombres) tiene lugar en hepatocitos y macrófagos del hígado, médula ósea, bazo y músculo [ver figura 3]. La pequeña cantidad de hierro en transporte (alrededor de 0.2 mg/kg) en el plasma y el líquido extracelular está unido a la proteína transferrina, que lleva el hierro para satisfacer las necesidades tisulares en todo el organismo.7,8
El eritrón se define como la totalidad de elementos eritroides en todas las fases de desarrollo y los tejidos en los que se producen.1 La mayoría del hierro en el organismo se localiza en el eritrón, y la mayoría del flujo diario de hierro pasa a través de este compartimiento. La vía predominante de flujo interno de hierro es unidireccional de la transferrina del plasma al eritrón, al sistema monocito-macrófago y después de regreso a la transferrina.
El flujo de hierro hacia y desde el eritrón constituye alrededor de las cuatro quintas partes del hierro que pasa a través del compartimiento de transferrina cada día. El reciclamiento eficaz del hierro de los eritrocitos recién destruidos o formados es una característica distintiva del metabolismo humano del hierro.
En condiciones fisológicas, menos del 0.05 porciento del hierro corporal total se adquiere o pierde cada día. La eficacia con la que el hierro se recicla en el organismo es única, ningún otro animal conserva el hierro en forma tan eficaz. Fisiológicamente, el eritrón adquiere hierro de transferrina a través de un receptor específico de transferrina localizado en la superficie de la membrana de las células eritroides inmaduras. La mayoría de este hierro se usa para la síntesis de hemoglobina y después entra a la circulación en los eritrocitos. Pequeñas cantidades de hierro se almacenan en la ferritina (la proteína almacenadora de hierro), entran a enzimas que contienen hierro de las células eritroides inmaduras o se pierden en los productos de la eritropoyesis ineficaz.
Los eritrocitos viejos son fagocitados por macrófagos especializados en el bazo, la médula ósea y el hque regresan la mayoría del hierro eritroide catabolizado al compartimiento de transferrina, donde comienza su ciclo de nuevo. La fagocitosis de las células eritroides inmaduras y de los eritrocitos viejos explica la mayoría de todo el hierro almacenado que se encuentra en los macrófagos del hígado, médula ósea y bazo, ya que nada o casi nada de este hierro deriva de la transferrina.
Por el contrario, las células parenquimatosas del hígado pueden captar hierro de la transferrina o entregarlo a la misma. La quinta parte restante del hierro que se moviliza hacia y desde la transferrina corresponde a cantidades más o menos iguales de hierro absorbido y perdido, que se desplaza de los compartimientos de transferrina del plasma y extravascular, se mueve hacia y desde los hepatocitos y se intercambia entre la transferrina extravascular y los tejidos parenquimatosos.
BALANCE DEL HIERRO
El balance del hierro es el resultado de la diferencia entre la cantidad de hierro que recibe y que pierde el organismo. Debido a que los humanos son incapaces de excretar el exceso de hierro, el balance está regulado fisiológicamente por el control de la absorción.9 Los dos principales factores que influyen en la absorción del hierro son el nivel de las reservas corporales de hierro y la magnitud de la eritropoyesis. Si la reserva de hierro aumenta la absorción disminuye y viceversa. La absorción aumenta al aumentar la actividad eritropoyética, en especial con la eritropoyesis ineficaz. El hierro de la dieta se absorbe principalmente por las células mucosas del duodeno y la porción superior del yeyuno. La base molecular para la absorción de hierro por las células mucosas de la porción superior del intestino delgado sigue siendo incierta, pero existen evidencias de un mecanismo mucina-mobilferrina-integrina.10
TRANSPORTE, ALMACEN Y REGULACION DEL HIERRO
Desde el punto de vista fisiológico, tres proteínas (transferrina, receptor de transferrina y ferritina) son responsables del transporte y almacén del hierro. A su vez, la expresión coordinada de estas tres proteínas está controlada por una cuarta clase de proteínas, las proteínas reguladoras del hierro (PRH). La transferrina es la proteína transportadora que proporciona el único medio fisiológico para el transporte de este elemento a las células. El receptor de transferrina es el receptor de la superficie celular que permite la entrada del complejo transferrina-hierro a las células. La ferritina, la proteína de almacén, proporciona un reservorio en la célula en el que puede depositarse el hierro hasta que se requiera para el metabolismo. Dentro de las células, la expresión del receptor de transferrina y de la ferritina está regulado por un ácido ribonucleico mensajero (ARNm) de proteínas de unión identificadas actualmente como proteínas reguladoras del hierro. Cada una de las proteínas principales ha sido aislada, purificada y caracterizada [ver figura 4].
TRANSFERRINA
La transferrina [ver figura 4] transporta el hierro a los eritrocitos en desarrollo y a otras células del organismo, en especial las que están en crecimiento o proliferación.11,12 En los humanos el hepatocito es la fuente de casi toda la apotransferrina circulante, que se divide en porciones más o menos iguales entre el plasma y los líquidos extravasculares.
La apotransferrina es una glucoproteína transportadora con vida media de alrededor de ocho días,11 después de entregar el hierro a las células, la apoferritina retorna con rapidez al plasma para funcionar de nuevo como transportadora. La apotransferrina es una glucoproteína de una sola cadena bilobular con dos lóbulos semejantes en los extremos N- y C-terminales.13 Debido a que un solo ión férrico puede unirse en cada lóbulo, existen cuatro formas de la molécula: (1) apotransferrina, (2) transferrina monomérica, con un átomo de hierro unido al lóbulo N-terminal (FeNTf), (3) transferrina monoférrica, con un átomo de hierro unido al lóbulo C-terminal (FecTf), y (4) transferrina diférrica (Fe2Tf). La saturación de la transferrina es la proporción de sitios disponibles para unión de hierro en la transferrina que están ocupados por átomos de hierro, expresado en porcentaje. Las proporciones relativas de apotransferrina, transferrina monoférrica y transferrina diférrica son una función estocástica de la saturación de transferrina.
RECEPTOR DE TRANSFERRINA
El receptor de transferrina [ver figura 4] es la única vía de entrada fisiológica a la célula para el hierro unido a transferrina, y también ayuda a liberar el hierro de la transferrina para su uso en la célula.14,15 Los receptores de transferrina se localizan en la superficie de la membrana de todas las células nucleadas, en números que dependen de los requerimientos celulares de hierro. Para cada célula determinada, el determinante predominante del aporte de hierro a la célula es el número de receptores de transferrina expresados en la superficie de la célula. Por ello, el número de receptores de transferrina es más alto en las células de la médula eritroide, el hígado y la placenta. Desde el punto de vista estructural, el receptor de transferrina consiste en dos subunidades transmembrana glucoproteicas idénticas unidas por un puente disulfuro [ver figura 4]. La eficacia con la que el receptor de transferrina puede entregar el hierro unido a la transferrina depende del contenido de hierro de la transferrina. Cada subunidad puede fijar una molécula de transferrina, de modo que el receptor puede aceptar dos moléculas. El receptor dimérico puede entonces permitir la entrada de cuatro átomos de hierro si cada transferrina es diférrica o solo dos átomos de hierro si cada una es monoférrica. Tanto el pH como el contenido de transferrina determinan la magnitud de la afinidad del receptor de transferrina por la transferrina. A un pH fisiológico de 7.4, con suficiente transferrina con hierro para saturar los receptores disponibles, el receptor de transferrina casi no tiene afinidad por la apotransferrina, afinidad moderada por la transferrina monoférrica y la mayor afinidad por la transferrina diférrica (más de cuatro veces mayor que para la transferrina monoférrica).11 Al disminuirse el pH a alrededor de 5, la afinidad del receptor de transferrina por la apotransferrina aumenta a una magnitud equiparable a la que tiene por la transferrina diférrica a un pH de 7.4.
FERRITINA
La ferritina [ver figura 4] es la principal proteína para almacén de hierro y tiene una vida media de alrededor de 60 horas (para ferritina hepática). Casi todas las células contienen ferritina, que funciona tanto como un sitio de almacén seguro para hierro como una reserva accesible que ha sido adquirida por la célula ante la existencia en exceso a sus necesidades metabólicas inmediatas. En consecuencia, las mayores cantidades de ferritina se encuentran en las células dedicadas al almacén de hierro (i.e., macrófagos y hepatocitos) y en las células con los mayores requerimientos para la síntesis de compuestos que contienen hierro (i.e., células eritroides en desarrollo).16 La apoferritina, que tiene un peso molecular de 440,000, es una concha esférica con radio de alrededor de 6 mm, cada molécula puede almacenar hasta 4,500 átomos de hierro en el interior en un centro polinuclear de hidrofosfato férrico. La proteína de apoferritina está formada por 24 subunidades oblongas que se denominan como H (pesadas, con un peso molecular de 21,100 daltons) y L (o ligeras, con un peso molecular de 19,700 daltons). Las subunidades tienen homología considerable, pero existe un sitio de ceruloplasmina en la subunidad H17 y no en la subunidad L. La abundancia relativa de subunidades H y L está determinada tanto por el tipo celular específico como por el estado férrico de la célula. El mayor número de subunidades L se encuentra en las células de tejidos que tienen función especializada de almacén de hierro, como el hígado y el bazo,18 mientras que existen más subunidades H en células de tejidos que no funcionan como depósitos de hierro, como el músculo cardiaco. Para un tipo específico de célula, se encuentra mayor proporción de subunidades L cuando es mayor el almacén de hierro. Estas observaciones sugieren que las ferritinas con mayor proporción de subunidades H son más activas en el metabolismo y que las que tienen mayor abundancia de subunidades L sirven para almacenar el hierro a más largo plazo.
PROTEINAS REGULADORAS DE HIERRO
Las proteínas críticas que permiten al hierro autorregular su disponibilidad intracelular son las proteínas reguladoras del hierro PRH1 y PRH2 [ver figura 4]. Las proteínas reguladoras del hierro son proteínas que unen ARN citoplásmico que funcionan en una forma de acción-trans sobre los ARNm que contienen un elemento que responde al hierro (ERH), una estructura reguladora de acción-cis.19,20 El ERH, que es el sitio de unión para las PRH, es una estructura en forma de pasador compuesta de una secuencia conservada de 28 nucleótidos. La región 3í no traducida del ARNm del receptor de transferrina, la región 5í no traducida del ARNm de la ferritina y el ARNm de la forma eritroide específica de la sintetasa de ácido d-aminolevulínico (AAL) contienen ERH.20 Los tejidos que contienen PRH muestran diferencias importantes en la cantidad de PRH1 y PRH2. Los tejidos con la mayor cantidad de almacén de hierro, como el hígado y el bazo, tienen mayor cantidad de PRH1.21 Los tejidos con mayor utilización del hierro, como el corazón y el músculo esquelético, contienen la mayor cantidad de la PRH2.22
Se ha encontrado que la PRH1 es la aconitasa citosólica23 (hidro-liasa de citrato [isocitrato]), una proteína con sulfuro de hierro que actúa enzimáticamente dentro del ciclo de Krebs en la interconversión de citrato a isocitrato. Dentro de la clula, la aconitasa existe en una forma citoplásmica (codificada por un gen en el cromosoma 9) y mitocondrial (codificada por un gen en el cromosoma 22). Tanto las formas citoplásmica como mitocondrial son enzimáticamente activas como enzima aconitasa, pero solo la forma citoplásmica puede funcionar como PRH. Un sitio específico (4Fe-4S) forma el sitio catalíticamente activo de la enzima aconitasa, en el que tres átomos de hierro se unen a la estructura de la proteína y el cuarto átomo se une a un azufre inorgánico en el sitio específico. Los cambios en la disponibilidad de hierro intracelular parecen alterar la conformación del sitio Fe-S, transformando de modo reversible la aconitasa enzimáticamente activa, que normalmente es incapaz de unirse al ARN con gran afinidad, en una forma que tiene gran afinidad de unión al ARN y que no tiene actividad de aconitasa. Estas conversiones permiten a la PRH1 monitorizar los cambios en la disponibilidad del hierro intracelular. La mayor cantidad de hierro intracelular aumenta las cantidades de PRH1 en la forma de baja afinidad de unión al ARN, disminuyendo la unión de la PRH1 al ERH. La reducción en el hierro intracelular disponible aumenta la cantidad de PRH1 en la forma de alta afinidad de unión al ARN, aumentando la unión de la PRH1 al ERH.
La PRH2 no tiene actividad de aconitasa, pero parece unirse al ERH con una afinidad indistinguible de la de la PRH1 y funcionar con la misma eficacia como represor de la traducción in vitro.22 La reducción en la disponibilidad de hierro intracelular aumenta la actividad de unión al ARNm de ambas PRH, pero por diferentes motivos.24 El control de la PRH1 es principalmente postraducción, y el cambio de la forma de no unión (con actividad de aconitasa) a la forma de unión al PRH (sin actividad de aconitasa) produce poco cambio en la cantidad de PRH1. Por el contrario, la mayor actividad de unión al ARNm de la PRH2 es el resultado de síntesis de novo.25 Las consecuencias de las diferencias en la distribución tisular y en los mecanismos de regulación del hierro de las PRH1 y PRH2 aún no se conocen, pero la dos PRH parecen funcionar de modo semejante como sensores de la disponibilidad de hierro intracelular.26
APORTE Y ALMACEN DE HIERRO EN LA CELULA
La transferrina, el receptor de transferrina y la ferritina actúan juntos para proporcionar hierro a la célula [ver figura 5].11 Al inicio dos moléculas de transferrina (monoférrica o diférrica) se unen a un receptor de transferrina sobre la superficie celular. Con la eritropoyesis normal, la mayoría del aporte de hierro al eritrón es proporcionado por transferrina diférrica cuando la saturación de transferrina es de 33 porciento o mayor. Con saturaciones de transferrina de 19 porciento o menos la mayoría del hierro proviene de transferrina monoférrica. Ambas formas de transferrina aportan hierro a una saturación de 20 a 32 porciento. El complejo hierro-transferrina-receptor de transferrina se agrega a otros complejos en un orificio cubierto por clatrin?, que después es translocado al interior de la célula como una vesícula cubierta. Después de que el clatrin? es separado, la vesícula sin cubierta se fusionan con otras vesículas para formar un endosoma multivesicular. El endosoma se acidifica hasta un pH de 5.6 por una bomba de protones mientras viaja en el interior de la célula. Con la caída en el pH, el receptor de transferrina sufre cambios de conformación que ayudan a liberar el hierro de la transferrina mono o diférrica. Aunque aún no se conocen los mecanismos moleculares que participan, el hierro disociado es entonces transportado a través de la membrana endosómica para ser utilizado en la síntesis de compuestos que contengan hierro o para su almacén en la ferritina citoplásmica. En el endosoma a un pH ácido, la apotransferrina sin hierro se une en forma ávida al receptor de transferrina, y el complejo es transportado de regreso a la membrana celular en el endosoma. Aunque algunos endosomas se desvían hacia el aparato de Golgi para su resialilación y otros son reparados, la mayoría de los endosomas llegan en forma directa a la superficie celular y se fusionan con la membrana. El complejo apotransferrina-receptor de transferrina se expone entonces al pH de 7.4 del plasma, lo que causa pérdida de la afinidad de la apotransferrina por el receptor. La apotransferrina entra al reservorio circulante en el plasma y el receptor de transferrina está de nuevo disponible para unir a una transferrina que lleve hierro.11
REGULACION DE LA CAPTACION DE HIERRO EN LA CELULA Y SU ALMACEN POR PROTEINAS REGULADORAS DE HIERRO
Las PRH controlan la captación intracelular y el almacén del hierro al controlar la traducción en la síntesis del receptor de transferrina y de la ferritina.19,20 Las PRH1 y PRH2 responden en forma semejante a cambios en el hierro disponible dentro de la célula y parecen fijarse al ERH con igual avidez y eficacia.26 La síntesis del receptor de transferrina se regula al controlar la estabilidad del ARNm citoplásmico del receptor de transferrina. Existen tres ERH en la región no traducida 3í (RNT3í) del ARNm. Al unirse la PRH al ERH en la RNT3í, la estabilidad del ARNm aumenta y se reduce su degradación citoplásmica. La cantidad de ARNm citoplásmico del receptor de transferrina aumenta, y la velocidad de síntesis del mismo también. Al incrementarse el número de receptores de transferrina en la superficie celular, la captación celular de hierro será mayor. La síntesis de ferritina esta regulada por un control en la traducción del ARNm de ferritina en el citoplasma. Existe un solo ERH en la región no traducida 5í (RNT5í) del ARNm de la ferritina. Cuando este ERH en la RNT5í se une a una PRH, se detiene la traducción del ARNm de la ferritina. Al producirse menos ferritina disminuye el almacén de hierrro y aumenta la disponibilidad de hierro celular. Por lo tanto, los cambios en la cantidad de la PRH tienen efectos opuestos en la producción del receptor de transferrina y la ferritina. Si cae el nivel de hierro intracelular, la cantidad de PRH de alta afinidad aumenta. A mayor cantidad de PRH la unión a los ERH en la RNT3í estabiliza al ARNm del receptor de transferrina, aumentando la producción del receptor de transferrina y la disponibilidad de hierro intracelular. El aumento en las PRH también disminuye la síntesis de ferritina al unirse a la RNT5í del ARNm , disminuyendo el almacén de hierro y aumentando la disponibilidad de hierro intracelular. Por el contrario, el aumento en el hierro intracelular disminuye la cantidad de PRH de alta afinidad, disminuyendo la captación de hierro por la célula al reducir la síntesis de la proteína del receptor de transferrina y aumenta el almacén de hierro al incrementar la síntesis de proteína ferritina. De esta forma coordinada, las PRH actúan para conservar la disponibilidad de hierro intracelular al controlar la traducción en la síntesis de las proteínas encargadas de la captación y almacenamiento del hierro.19,20
Evaluación de laboratorio del metabolismo del hierro
Se observan cambios característicos en la función, transporte y almacén de hierro al aumentar o disminuir su cantidad en el organismo [ver figura 6 y tabla 1]. Pueden usarse métodos tanto directos como indirectos para evaluar el estado corporal del hierro, pero ningún indicador o combinación de indicadores proporciona una forma óptima para evaluar al aporte y almacén del hierro en el organismo en todas las situaciones. Además, los métodos disponibles deben interpretarse tomando en cuenta como se afecta cada medida por la presencia concomitante de inflamación, infección, neoplasias, enfermedad hepática, deficiencia de ácido ascórbico o desnutrición.
MEDIDAS DIRECTAS DEL ESTADO DEL HIERRO CORPORAL
Las medidas directas del hierro corporal proporcionan la manera cuantitativa de evaluar a este elemento y tienen una gran sensibilidad y especificidad. La flebotomía cuantitativa permite calcular las reservar de hierro que pueden movilizarse para la producción de hemoglobina. Este método típicamente requiere la eliminación de alrededor de 500 ml de sangre una o dos veces por semana hasta que se agoten las reservas de hierro, lo que se indica por una reducción sostenida de la concentración de hemoglobina a menos de 10 g/dl sin que se continúen las flebotomías. El hierro almacenado y movilizado puede calcularse a partir de la cantidad de hierro de la hemoglobina extraído, corrigiendo por la diferencia entre las concentraciones inicial y final de hemoglobina y por la absorción de hierro de la dieta durante el periodo de flebotomía. La flebotomía cuantitativa puede ser útil para establecer el diagnóstico retrospectivo de hemocromatosis asociada al HLA en homocigotos y de otras formas de sobrecarga de hierro. Este método no puede usarse en la mayoría de las formas de anemia con exceso de hierro o en pacientes dependientes de transfusión, y no es útil en pacientes con reducción en la cantidad del hierro corporal. La aspiración y biopsia de médula ósea son de mayor utilidad en la evaluación de los pacientes con deficiencia de hierro u otras condiciones en las que se requiere evaluar el aporte de hierro para la eritropoyesis y las características morfológicas de la hematopoyesis. La tinción de la médula con azul de Prusia puede proporcionar información sobre (1) las reservas de hierro en los macrófagos al evaluar la cantidad de hemosiderina en la médula y (2) la morfología y proporción de sideroblastos en la médula (eritroblastos que contienen uno o mas gránulos positivos al azul de Prusia, que pueden identificarse por microscopía de luz). Si se requiere, puede medirse también el almacén de hierro en los macrófagos en una muestra de médula por medio de la determinación química del hierro no hem. El estudio de la médula ósea no proporciona datos sobre el grado de acúmulo de hierro en las células parenquimatosas, por lo que no suele ser útil en la evaluación de la sobrecarga de hierro. La biopsia hepática con determinación cuantitativa de hierro y estudio histoquímico es el mejor método para evaluar el hierro corporal en pacientes con todas las formas de sobrecarga sistémica de hierro.27 En este grupo de pacientes, la determinación cuantitativa de la concentración de hierro no hem en el tejido hepático obtenido por biopsia es el método de referencia para la medición del acúmulo de hierro en el organismo. El examen del patrón de depósito de hierro en los hepatocitos y las células de Kupffer en la muestra de biopsia proporciona información sobre la magnitud relativa del acúmulo de hierro en las células parenquimatosas y los macrófagos. Aunque estos métodos directos para evaluar el estado del hierro pueden proporcionar resultados definitivos en las circunstancias apropiadas, todos son métodos invasivos que pueden ser inaceptables, incómodos y, en el caso de la biopsia hepática, riesgosos para los pacientes. Se han examinado varias alternativas no invasivas, incluyendo la determinación de la susceptibilidad magnética hepática,28-30 la tomografía computada,31 y la imagen por resonancia magnética,32 pero en la actualidad estos procedimientos están en fase de investigación.
CALCULO INDIRECTO DEL ESTADO DEL HIERRO CORPORAL
Comparados con los métodos directos de medición del hierro corporal, los métodos indirectos son menos cuantitativos y no son tan sensibles ni tan específicos porque todos los métodos indirectos se afectan por diversas condiciones. A pesar de estas limitantes, los indicadores indirectos son accesibles y requieren solo una muestra de sangre u orina. La concentración de ferritina en plasma es la medición más empleada como indicador indirecto de las reservas corporales de hierro.33 En personas sanas, las concentraciones de ferritina en plasma suelen disminuir al disminuir la reserva de hierro y aumentan cuando éste se acumula [ver figura 6 y tabla 1]. Por el contrario, en personas con diversos padecimientos, la relación entre el hierro en plasma y el corporal puede estar muy alterado.34 Por ejemplo, la deficiencia de ácido ascórbico puede disminuir la concentración de ferritina en plasma sin que cambien las reservas corporales de hierro. En ausencia de deficiencia de ácido ascórbico, y quizá de hipotiroidismo y desnutrición severa, una concentración de ferritina en plasma de menos de 12 µg/L es muy específica de deficiencia de hierro. Las concentraciones de ferritina en plasma que son normales o aumentadas pueden ser difíciles de interpretar porque la ferritina plasmática es un reactante de fase aguda, lo que significa que su síntesis y concentración plasmática aumenta con la infección, la inflamación, la enfermedad hepática aguda y crónica y las neoplasias. En algunos casos estos trastornos también provocan liberación de ferritina de células dañadas (en especial con hepatopatía o quimioterapia citotóxica) o interfieren con la depuración de la ferritina circulante. Este aumento en la ferritina del plasma puede ocultar la deficiencia de hierro o sugerir en forma falsa exceso del mismo. Se ha sugerido que la concentración máxima de ferritina en plasma en respuesta a un aumento en el hierro corporal es de alrededor de 4,000 µg/L, lo que quizá representa el límite fisiológico superior de síntesis.35 Pueden presentarse concentraciones por arriba de 4,000 µg/L por la liberación de ferritina intracelular de células dañadas. La ferritina plasmática puede aumentar también por hemólisis y eritropoyesis ineficaz sin ningún cambio en las reservas corporales de hierro. Por último, también se encuentran concentraciones elevadas de ferritina sin sobrecarga de hierro en el recién reconocido síndrome de hieperferritinemia hereditaria y el de catarata congénita autosómico dominante.36
La saturación de transferrina es el indicador indirecto más útil para investigar hemocromatosis ligada al HLA y otras formas de sobrecarga parenquimatosa de hierro27 [ver figura 6 y tabla 1]. Los valores de saturación de transferrina por encima del 50 porciento para las mujeres y del 60 porciento para los hombres requieren de mayor investigación. Aunque útil como prueba de escrutinio, no refleja en forma cuantitativa las reservas de hierro del organismo. La labilidad de la saturación de transferrina, en especial en la hemocromatosis ligada al HLA temprana, posee otros problemas de interpretación. La saturación de transferrina también se altera por infección, inflamación, enfermedad hepática, neoplasias, quimioterapia citotóxica y otros trastornos. Por lo tanto, existen condiciones, como la enfermedad hepática, en las que la saturación de ferritina sérica y la saturación de transferrina pueden estar elevadas sin un aumento en la reserva de hierro del organismo. La saturación de transferrina disminuye tanto en la deficiencia de hierro [ver figura 6 y tabla 1] como en la anemia por inflamación crónica,37 pero los valores de saturación de transferrina se superponen mucho en estas dos condiciones, lo que hace la medición de poca utilidad para distinguir entre ambos padecimientos.
La concentración del receptor de transferrina en plasma es una prueba nueva útil para detectar deficiencia de hierro en presencia de inflamación o infección.38 El receptor de transferrina que se encuentra en el plasma circulante es una forma trucada del receptor de transferrina celular dimérico. El receptor de transferrina soluble corresponde al dominio citoplásmico N-terminal, que al parecer se desprende por acción proteolítica de la membrana celular. Se considera que la concentración del receptor de transferrina en plasma depende de la masa total del receptor tisular. En personas normales la mayoría de los receptores celulares de transferrina se encuentran en los precursores eritroides en la médula ósea, y la mayoría de los receptores solubles provienen de estas células eritroides. En consecuencia, la concentración del receptor de transferrina en plasma está determinada principalmente por la actividad de la médula eritroide. La concentración plasmática del receptor de transferrina está disminuida cuando existe hipoplasia (o aplasia) eritroide, como en el caso de la insuficiencia renal crónica o la anemia aplástica. Por el contrario, la concentración del receptor de transferrina en plasma aumenta en pacientes con hiperplasia eritroide, esto es, en pacientes con talasemia mayor, anemia de células falciformes y otros estados hemolíticos crónicos. En ausencia de hiperplasia eritroide, un aumento en la concentración del receptor de transferrina proporciona un método útil para detectar la deficiencia de hierro tisular. La concentración del receptor de transferrina puede ser especialmente útil para distinguir la anemia por deficiencia de hierro de la anemia asociada con la inflamación crónica o la infección. Mientras que la concentración sérica de ferritina puede aumentar con niveles normales, disminuidos o ausentes de hierro en los pacientes con infección, inflamación o enfermedad hepática, la concentración del receptor de transferrina en plasma parece no afectarse por estos trastornos concomitantes. En estas circunstancias, la concentración del receptor de transferrina en plasma proporciona un indicador nuevo y aparentemente más confiable de la deficiencia de hierro.39 La concentración del receptor de transferrina en plasma no es útil para diagnosticar sobrecarga de hierro.
La excreción urinaria de hierro con agentes quelantes, como la desferroxamina o el ácido dietilenetriamino penta-acético (DPTA), constituye otro indicador del estado del hierro en el organismo,7 que es especialmente útil para evaluar la sobrecarga de hierro. Después de administrar una dosis estándar de un agente quelante, existe considerable superposición de las tasas de excreción de hierro en la orina en las personas con reservas normales y en las que tienen reservas disminuidas, de modo que esta prueba no es útil para diagnosticar deficiencia de hierro. La medición de la excreción de hierro en orina en ocasiones puede ayudar a detectar la sobrecarga del mismo, pero este indicador puede verse influenciado por la deficiencia de ácido ascórbico, infecciones, inflamación, enfermedad hepática y otros padecimientos.
La protoporfirina zinc eritrocitaria es un indicador de la cantidad de hierro disponible en la médula ósea cuando se produjeron los eritrocitos circulantes.39 El paso final en la biosíntesis del hem consiste en la inserción de un ión ferroso a una protoporfirina IX para producir el hem. En ausencia de hierro se inserta zinc, lo que origina una protoporfirina zinc, que persiste durante toda la vida del eritrocito. Como resultado, la concentración de protoporfirina zinc refleja el aporte de hierro a la médula eritroide durante las semanas previas. Como con algunos otros indicadores del estado del hierro, el aumento en la protoporfirina zinc eritrocitaria no tiene especificidad y es influenciada por varias condiciones que limitan el aporte de hierro, como la deficiencia de ácido ascórbico, la infección, la inflamación, la enfermedad hepática y las neoplasias. Los niveles de protoporfirina zinc también aumentan en muchas anemias sideroblásticas y, en especial, en la intoxicación crónica por plomo. La protoporfirina zinc eritrocitaria no es de utilidad en el diagnóstico de la sobrecarga de hierro, pero es útil como prueba de escrutinio para la intoxicación por plomo.
El examen de la sangre periférica no ayuda a detectar sobrecarga de hierro, que no produce alteraciones diagnósticas, y puede detectar cambios en la reducción del hierro corporal solo después de que la depleción de las reservas de hierro restringe el aporte de hierro para la eritropoyesis. Estas limitaciones se aplican cuando el frotis es examinado en forma visual o con el uso de instrumentos para determinar concentración de hemoglobina, hematocrito, índices eritrocitarios, distribución del volumen de los eritrocitos y cuenta de reticulocitos.8 Los cambios observados en la sangre periférica no son específicos de deficiencia de hierro y pueden ser también causados por cualquier trastorno que afecte la síntesis de hemoglobina, como la talasemia, la infección, la inflamación y las neoplasias.
Exceso de hierro
El exceso de hierro se origina de un aumento constante en el aporte de hierro en relación con los requerimientos, y causa cambios característicos en la función, transporte y almacén del hierro [ver figura 6 y tabla 1]. La cantidad de hierro corporal suele controlarse en condiciones normales por regulación en la absorción dietética de hierro. Se desarrolla exceso de hierro en condiciones que modifican o superan la regulación de la absorción del ión en el intestino [ver tabla 2]. Debido a que los humanos no tienen una forma fisiológica de eliminar el exceso de hierro, cualquier incremento persistente en la ingesta puede eventualmente causar exceso de hierro. Independientemente de la causa del acúmulo del hierro, cuando la cantidad del mismo excede la capacidad del organismo para secuestrar el exceso se desarrolla daño tisular potencialmente letal. Las características precisas de las consecuencias patológicas de la sobrecarga de hierro son resultado, en parte, de la magnitud de la carga de hierro corporal, la velocidad a la que ha ocurrido el aumento, la distribución específica del exceso del ión entre los sitios de almacén en macrófagos y los depósitos, potencialmente más dañinos, en células parenquimatosas, y la coexistencia de condiciones que pueden disminuir (v.gr., deficiencia de ácido ascórbico) o pueden empeorar (v.gr., uso de alcohol o hepatitis) la evolución. Las consecuencias más comunes de la sobrecarga de hierro incluyen enfermedad hepática, enfermedad pancreática asociada con diabetes mellitus, trastornos endócrinos asociados con insuficiencia gonadal, disfunción cardiaca, artropatía y, en ocasiones, alteraciones neurológicas y psicológicas.
EXCESO DE HIERRO CAUSADO POR MAYOR ABSORCION
Puede ocurrir mayor absorción de hierro por un control anormal en su absorción aún cuando la dieta contenga cantidades normales de hierro biodisponible, como en la hemocromatosis hereditaria (asociada al HLA), o por una dieta con tanto exceso de hierro biodisponible que supera los mecanismos de control, como en el exceso dietético de hierro que ocurre en africanos.
Hemocromatosis hereditaria (asociada al HLA)
La hemocromatosis hereditaria es la causa más común de sobrecarga de hierro en los Estados Unidos. Hasta el 10 porciento de la población es heterocigota para esta condición, y se cree que el estado homocigoto afecta hasta al 0.5 porciento de la población, o alrededor de un millón de personas.27,40 En este trastorno el defecto genético depende de una alteración en el control de la absorción de hierro que causa un aumento inapropiado en la captación del mismo, con incremento progresivo en las reservas corporales. El exceso de hierro tiene un patrón de depósito principalmente parenquimatoso al inicio, sobre todo en los hepatocitos, y subsecuentemente el hierro se acumula en páncreas, corazón y otros órganos.27 Es característico que el hierro de los macrófagos en la médula ósea puede ser normal o incluso estar disminuido a pesar de un depósito parenquimatoso importante [ver figura 6 y tabla 1]. En los homocigotos que presentan hemocromatosis ligada al HLA en la edad media o después, la tétrada clásica de signos clínicos consiste en enfermedad hepática, diabetes mellitus, pigmentación cutánea y falla gonadal.41 Se desarrolla insuficiencia cardiaca en alrededor del 10 a 15 porciento de los homocigotos no tratados. Con menos frecuencia, los pacientes más jóvenes con hemocromatosis presentan alteraciones cardiacas y falla gonadal como signos clínicos iniciales.42 Las reservas de hierro corporal suelen haber aumentado de la cantidad normal de 1 g o menos a 15 o 20 g o más cuando ocurren los síntomas de daño parenquimatoso, por lo general en la edad media o tardía.43 Los aumentos adicionales en el hierro corporal pueden ser fatales, aunque algunos pacientes son capaces de tolerar una reserva total de hierro de hasta 40 a 50 g. Los factores ambientales, incluyendo el contenido de hierro de la dieta y el uso de alcohol, pueden influir mucho en la velocidad y severidad del daño orgánico.44 En los heterocigotos la hemocromatosis ligada al HLA se expresa en forma incompleta, y solo alrededor del 25 porciento de estas personas sufren un aumento leve y aparentemente inofensivo en las reservas de hierro, con cantidades totales de 4 o 5 g.29 Se ha reportado un exceso más importante de hierro en los pacientes heterocigotos para hemocromatosis con esferocitosis hereditaria coexistente, pero el efecto de un alelo para hemocromatosis sobre la reserva de hierro es incierta en la anemia sideroblástica idiopática y en otros trastornos hereditarios o adquiridos.
La detección de la hemocromatosis ligada al HLA aún depende de la expresión fenotípica de la enfermedad. Aún no se ha identificado el gen responsable, y el análisis directo del ADN genómico no es factible como medio para identificar a los homocigotos. El locus de la hemocromatosis está unido en forma estrecha al locus del HLA-A en el brazo corto del cromosoma 6. Por lo tanto, si se identifica un individuo afectado (probando), puede usarse la tipificación del HLA para clasificar los genotipos de los familiares en primer grado como normales, heterocigotos u homocigotos. En cambio, la tipificación del HLA no es útil como instrumento de escrutinio de población porque los haplotipos HLA específicos ligados al locus de la hemocromatosis pueden ser diferentes en cada familia. Para propósitos de escrutinio, la medición del hierro en plasma, la saturación de transferrina y la ferritina en plasma27 proporcionan el mejor medio indirecto para estudiar a personas que son homocigotas para la hemocromatosis ligada al HLA, aunque debe reconocerse que la ferritina en plasma puede ser normal en un pequeño número de pacientes con hemocromatosis ligada al HLA.45 Si cualquiera de estas medidas es anormal, está indicada una evaluación posterior. La biopsia hepática permite un diagnóstico definido de la hemocromatosis ligada al HLA.27 La evaluación debe incluir determinación cuantitativa de la concentración de hierro no hem, evaluación histoquímica del patrón de depósito de hierro y evaluación patológica de la lesión tisular. El cálculo del índice hepático de hierro (la concentración hepática de hierro [expresada como µmol Fe/g de tejido hepático en peso seco] dividida entre la edad del paciente [expresada en años])46 puede ser útil para distinguir a los homocigotos para la hemocromatosis asociada al HLA de los heterocigotos o de los pacientes con aumento en el hierro corporal asociado con enfermedad hepática crónica (por lo general alcohólica). Es típico encontrar aumento en el índice de hierro hepático a 2.0 o más en las personas con hemocromatosis asociada al HLA homocigotas, mientras que el índice suele ser menor de 1.8 en los heterocigotos y menos de 1.4 en los pacientes con hepatopatía alcohólica crónica. En ciertos casos puede usarse la flebotomía cuantitativa para establecer el diagnóstico de estado homocigoto en forma retrospectiva si se eliminan más de 5 g de hierro. Siempre que se establezca el diagnóstico de hemocromatosis es obligatorio evaluar a los familiares con riesgo de la enfermedad. El escrutinio debe incluir no solo a los hermanos, sino también a los padres e hijos por la posibilidad de parejas homocigotas-heterocigotas. En estos estudios familiares puede ser útil la tipificación del HLA.
Tratamiento de la hemocromatosis asociada al HLA El tratamiento de elección para la hemocromatosis asociada al HLA es la flebotomía para reducir y mantener el hierro corporal en niveles normales o casi normales. En los pacientes con hemocromatosis asociada al HLA e insuficiencia cardiaca se ha sugerido el uso de flebotomía y tratamiento quelante. El tratamiento con flebotomías debe iniciarse en cuanto se hace el diagnóstico de estado homocigoto para la enfermedad, su postergación solo aumenta el riesgo de daño orgánico por exceso de hierro. El programa de flebotomía debe eliminar 500 ml de sangre (conteniendo 200 a 250 mg de hierro) una vez por semana o, en pacientes con exceso importante, dos veces por semana,47 hasta que el paciente tenga deficiencia de hierro. Antes de cada flebotomía debe medirse el hematocrito o la hemoglobina. Al principio estos deben disminuir alrededor de 10 porciento de sus valores iniciales, pero después pueden aumentar al aumentar la velocidad de la eritropoyesis para satisfacer las demandas de la flebotomía. Debe medirse en forma regular la ferritina, el hierro y la saturación de transferrina, para vigilar el progreso en la eliminación del hierro. Al eliminar hierro la concentración de ferritina en plasma disminuirá en forma progresiva, pero el hierro y la saturación de transferrina permanecerán elevados hasta que las reservas estén casi agotadas. Por último, cuando se ha eliminado todo el hierro acumulado la ferritina disminuirá a menos de 12 µg/L, el hierro y la saturación de transferrina caerán y la concentración de hemoglobina disminuirá a menos de 10 g/dl durante dos semanas sin otra flebotomía. Para ilustrar qué tanto tiempo puede requerirse el tratamiento, si la reserva inicial de hierro es de 25 g, su eliminación total con flebotomías semanales puede necesitar de dos años o más. Después de haber terminado la eliminación del hierro se requiere un programa de por vida de flebotomías de mantenimiento para evitar su reacúmulo. Típicamente se necesitan flebotomías de 500 ml cada tres a cuatro meses. El objetivo de la fletobomía de mantenimiento debe ser mantener una saturación de transferrina normal con una concentración de ferritina en plasma de menos de alrededor de 50 µg/L.
La concentración de hierro no hem hepático es un factor de riesgo importante para el desarrollo de cirrosis en la hemocromatosis asociada al HLA. En ausencia de hepatopatía alcohólica, no suele desarrollarse fibrosis o cirrosis hasta que el hierro almacenado en el hígado alcanza una concentración de 400 µmol Fe/g de hígado en peso seco (rango normal: alrededor de 3 a 30 µmol Fe/g de hígado en peso seco).46 Si el tratamiento con flebotomía elimina la carga de hierro antes que se desarrolle cirrosis la esperanza de vida para el paciente es normal.43 Si por el contrario ya existe cirrosis, el riesgo de carcinoma hepatocelular aumenta en más de 200 veces. En la hemocromatosis asociada al HLA se desarrollan hepatomas casi en forma exclusiva en los pacientes con cirrosis hepática, siendo la causa de muerte en el 20 a 30 porciento de estos pacientes, incluso después de la eliminación exitosa del exceso de hierro. En la actualidad la cirrosis y el carcinoma hepatocelular son las dos principales causas de muerte en la hemocromatosis asociada al HLA.48 Casi siempre está indicado el tratamiento con flebotomías para los pacientes con hemocromatosis asociada al HLA, incluso si ya existe cirrosis o daño orgánico, porque puede detenerse la progresión de la enfermedad e incluso mejorar parte de la disfunción orgánica.
Anemias con exceso de hierro
En los pacientes con el grupo heterogéneo de padecimientos clasificados como anemias con exceso de hierro, puede desarrollarse sobrecarga severa de hierro como resultado de una mayor absorción gastrointestinal del mismo.49 Cualquier transfusión de eritrocitos que reciban estos pacientes contribuye al acúmulo del hierro. Estas anemias por acúmulo de hierro pueden ser heredadas o adquiridas, pero todas parecen tener en común una importante hiperplasia eritroide con eritropoyesis ineficaz marcada. Las anemias con sobrecarga de hierro incluyen anemia diseritropoyética congénita, deficiencia de piruvato cinasa, talasemia mayor (anemia de Cooley) y talasemia intermedia, talasemia-§ con hemoglobina E, diversas formas de anemia sideroblástica, algunas anemias mielodispásicas y varios otros trastornos anémicos en los que se altera la incorporación del hierro a la hemoglobina. Debido a que la magnitud de la eritropoyesis ineficaz, y no la severidad de la anemia, parece determinar la velocidad de acúmulo de hierro, puede desarrollarse sobrecarga severa de hierro en pacientes con anemia solo leve. Las manifestaciones clínicas y la patología que puede desarrollarse en pacientes con anemias con exceso de hierro es semejante a las de la hemocromatosis asociada al HLA, incluyendo enfermedad hepática, diabetes mellitus, trastornos endócrinos y disfunción cardiaca.
Otras formas de exceso de hierro causadas por aumento en la absorción
Algunos pacientes con hepatopatía crónica, incluyendo los que tienen cirrosis alcohólica y con derivación portocava,46 pueden sufrir grados leves o moderados de exceso de hierro como resultado de mayor absorción del hierro de la dieta. Los mecanismos responsables de la mayor captación gastrointestinal de hierro no se han identificado, aunque se han propuesto como factores etiológicos a la eritropoyesis ineficaz y a la hiperferremia asociada con alteraciones sideroblásticas y del folato inducidas por alcohol.7 Las reservas corporales de hierro están aumentadas solo un poco, típicamente a 2 a 4 g. El depósito de hierro predomina en las células de Kupffer, más que en los hepatocitos, como en la hemocromatosis asociada al HLA. Como ya se mencionó, la distinción entre la sobrecarga de hierro en los pacientes con hepatopatía crónica y la de los homocigotos para la hemocromatosis asociada al HLA casi siempre puede hacerse por determinación cuantitativa de hierro en una muestra de hígado obtenida por biopsia calculando el índice de hierro hepático.
Los pacientes sintomáticos con porfiria cutánea tarda, una porfiria hepática, suelen tener un aumento modesto en el hierro corporal que casi siempre se debe a mayor absorción gastrointestinal. En la porfiria cutánea tarda el hígado produce porfirinas fotosensibilizantes que después circulan hacia la piel.8 La causa del exceso de hierro se desconoce, pero la eliminación del hierro por flebotomía o tratamiento quelante casi siempre causa mejoría clínica y bioquímica importante. Recientemente se ha reconocido a la aceruloplasminemia como un trastorno autosómico recesivo del metabolismo del hierro que causa acúmulo del mismo tanto en el hígado como en el cerebro, manifestándose como un padecimiento neurológico en las etapas tardías de la vida.50,51 Se han descubierto otros defectos congénitos raros asociados con sobrecarga de hierro. En los pacientes con atransferrinemia prácticamente no existe transferrina. La absorción gastrointestinal de hierro está aumentada, pero debido a que no existen mecanismos fisiológicos para su transporte, casi nada del hierro absorbido puede ser captado por las células eritroides en desarrollo. El hierro absorbido se acumula entonces en el hígado, páncreas, corazón, tiroides y riñones,52 con casi nada de hierro depositado en la médula ósea o el bazo. Se han encontrado depósitos marcado de hierro hepático y fibrosis asociados con un defecto congénito que parece consistir en una captación defectuosa del hierro por los precursores eritroides, pero la fisiopatología de este trastorno aún no se conoce.53
Se ha descrito exceso dietético de hierro en africanos en por lo menos nueve países en la región sub-Sahara de Africa en asociación con una ingesta muy abundante de hierro por una bebida fermentada de maíz preparada en casa en tambos de acero.7 El acúmulo de hierro puede ser tan importante como el encontrado en la hemocromatosis asociada al HLA, y puede desarrollarse enfermedad hepática (con cirrosis y hepatoma), daño pancreático (con diabetes mellitus), trastornos endócrinos y disfunción cardiaca. Aunque durante mucho tiempo se consideró que la mayor ingesta en la dieta era la única causa de la mayor absorción de hierro en este padecimiento, los análisis de familias han sugerido que puede participar un gen no asociado al HLA,54 que puede ser común en poblaciones con ancestros africanos. Se ha demostrado sobrecarga severa de hierro en pacientes de los Estados Unidos con ancestros africanos,55 pero la relación con el exceso de hierro dietético de los africanos aún es motivo de investigación.
Sin duda la ingestión medicinal de hierro puede contribuir al acúmulo del mismo en los pacientes con estos padecimientos, pero aún no se tiene certeza del grado en el que el hierro administrado por vía oral puede aumentar las reservas corporales de hierro en los individuos normales. Aunque algunos reportes de casos han descrito exceso de hierro en pacientes que toman hierro medicinal por periodos prolongados, no puede excluirse en estos pacientes la participación potencial de un alelo no reconocido para la hemocromatosis asociada al HLA, ya que el gen aún no se identifica.
EXCESO DE HIERRO POR TRANSFUSIONES Y ADMINISTRACION PARENTERAL
Un programa adecuado de transfusiones puede mantener la vida en pacientes con anemia severa crónica y refractaria, pero el tratamiento transfusional aislado produce acúmulo progresivo por el hierro contenido en los eritrocitos transfundidos.56 El acúmulo de hierro por transfusiones ocurre al inicio sobre todo en los macrófagos, seguido de redistribución a los tejidos parenquimatosos [ver figura 6]. En los pacientes con anemias congénitas severas, como la talasemia mayor y el síndrome de Blackfan-Diamond, las transfusiones regulares pueden prevenir la muerte por anemia en la infancia y permitir un crecimiento y desarrollo normal durante la niñez. Las anemias adquiridas dependientes de transfusión, como la anemia aplástica, la aplasia pura de eritrocitos, los trastornos hipoplásicos o mielodisplásicos y otros padecimientos, pueden causar exceso de hierro corporal. Si las anemias dependientes de transfusión incluyen hiperplasia eritroide con eritropoyesis ineficaz, puede agregarse mayor absorción gastrointestinal de hierro para empeorar el acúmulo. En estos casos debe minimizarse la captación de hierro de la dieta por medio de un programa adecuado de transfusiones. Aunque no dependen de transfusiones, los pacientes con trastornos falciformes, como la anemia de células falciformes y la talasemia-§ con células falciformes, pueden adquirir una cantidad considerable de hierro por transfusiones crónicas para prevenir eventos cerebrovasculares, crisis dolorosas y otras complicaciones recurrentes. Debido a que el humano no tiene sistemas fisiológicos para eliminar el exceso de hierro, el hierro contenido en los eritrocitos transfundidos se acumula en forma progresiva y llega a dañar al hígado, corazón, páncreas y otros órganos. Suele ocurrir la muerte por insuficiencia cardiaca.57 En los pacientes más jóvenes la carga de hierro causa falla en el crecimiento y, en la adolescencia, retraso o ausencia de la maduración sexual. El hierro medicinal parenteral puede aumentar el exceso de hierro en los pacientes con anemias microcíticas refractarias que son diagnosticados en forma equívoca como con deficiencia de hierro.
Tratamiento del exceso de hierro por transfusiones
Con cada unidad transfundida de eritrocitos se agregan a la reserva corporal alrededor de 200 a 250 mg de hierro. La mayoría de los pacientes dependientes de transfusión requieren de 200 a 300 ml/kg/año de sangre. Por ejemplo, un adulto de 70 kg requiere alrededor de dos a tres unidades de sangre cada tres o cuatro semanas, lo que agrega de 6 a 10 g de hierro por día. La severidad de la toxicidad por hierro parece relacionarse con la magnitud de la carga de hierro en el organismo. Casi todos los pacientes que han sido tratados solo con transfusión y han recibido 100 o más unidades de sangre58 (alrededor de 20 a 25 g de hierro) han desarrollado depósitos de hierro en el corazón, con frecuencia asociados con signos de daño hepático, pancreático y endócrino.
Para los pacientes que son dependientes de transfusión o tienen anemia severa, la única manera de superar la limitación fisiológica que evita la eliminación de hierro es con el tratamiento con un agente quelante capaz de formar complejos con el hierro y permitir su excreción. El único agente quelante disponible en la actualidad para uso clínico es un sideróforo bacteriano que comenzó a usarse hace tres décadas, la desferroxamina B, un ácido trihidroxámico producido por Streptomyces pilosus. En la actualidad existen estudios clínicos que han demostrado la eficacia de la quelación de hierro como un enfoque terapéutico a su exceso,59 demostrando que la quelación regular de este elemento puede disminuir su exceso corporal, reducir la disfunción orgánica y mejorar la supervivencia.30,60 Aunque en la actualidad se han reconocido diversos efectos adversos tóxicos, en especial con el tratamiento intensivo, la desferroxamina es un medicamento muy seguro, incluso cuando se ha usado practicamente de por vida en algunos pacientes.
El tratamiento de quelación del hierro debe comenzarse pronto para evitar el acúmulo de cantidades tóxicas de hierro en los tejidos susceptibles y para mantener las reservas corporales de hierro en concentraciones asociadas con un riesgo bajo de muerte temprana y complicaciones clínicas.30,60 Mientras más se retrase el tratamiento quelante mayor será el riesgo de toxicidad por hierro. Debido a que la desferroxamina se absorbe poco después de la administración oral, el medicamento debe administrarse por vía parenteral para ser activo. Además, la desferroxamina se elimina con tanta rapidez de la circulación que se requieren infusiones prolongadas para tener un efecto terapéutico.61 Como resultado, para que sea útil en los pacientes con exceso de hierro, debe administrarse por infusión subcutánea o intravenosa lenta durante nueve a 12 horas de cada día por lo menos durante cinco días de la semana. En los pacientes con talasemia mayor y otras anemias refractarias congénitas que son dependientes de transfusión desde la infancia temprana, es mejor comenzar el tratamiento quelante después de 10 a 20 transfusiones, por lo general alrededor de los tres años de edad, y administrar infusión subcutánea lenta en dosis no mayores de 25 mg/kg de peso/día para minimizar el riesgo de retraso en el crecimiento.62 En los pacientes ancianos y adultos con anemias refractarias adquiridas que requieren de transfusión regular y en los que tienen enfermedad de células falciformes que son transfundidos en forma crónica para prevenir complicaciones también parece prudente el tratamiento temprano, comenzando después de la transfusión de 10 a 20 unidades de sangre. La dosis habitual de desferroxamina en estos pacientes no debe ser mayor de 50 mg/kg de peso corporal/día, administrados en nueve a 12 horas por infusión subcutánea lenta por lo menos cinco días de la semana. En algunos pacientes que no son capaces de tolerar el dolor y molestia local de la infusión subcutánea o que requieren de la reducción rápida del exceso de hierro puede administrarse desferroxamina intravenosa a través de un reservorio implantable con acceso venoso.63 Puede ser difícil cumplir con estos esquemas casi diarios de infusión prolongada subcutánea o intravenosa, y la falta de constancia es el principal obstáculo para que el tratamiento sea eficaz.64 La administración de ácido ascórbico puede aumentar la excreción de hierro inducida por desferroxamina, pero se asocia con el riesgo de redistribución interna de hierro desde sitios de almacén relativamente benignos en los macrófagos hasta reservorios potencialmente tóxicos en las células parenquimatosas. Aunque la evidencia es anecdótica,65 las dosis grandes de ácido ascórbico deben considerarse como peligrosas en los pacientes con sobrecarga de hierro. Por lo general la desferroxamina es segura y no tóxica en el paciente con exceso de hierro, pero se han reportado complicaciones sistémicas, incluyendo reacciones anafilactoides alérgicas, complicaciones infecciosas, trastornos visuales y disfunción auditiva,66 y retraso del crecimiento.62 Como resultado, debe incluirse la evaluación regular de toxicidad medicamentosa en el tratamiento de cualquier paciente que reciba desferroxamina, incluyendo audiometrías anuales, examen de retina y evaluación del crecimiento en niños y adolescentes.62 El riesgo de muchas de estas complicaciones puede reducirse ajustando la dosis de desferroxamina según la magnitud del exceso de hierro.67 En la actualidad se están desarrollando agentes quelantes activos por vía oral, pero ninguno está clínicamente disponible aún.68,69
OTRAS FORMAS DE EXCESO DE HIERRO
Se ha detectado exceso de hierro perinatal en asociación con algunas alteraciones metabólicas raras del neonato, incluyendo tirosinemia hereditaria (hipermetioninemia), síndrome cerebrohepatorrenal o síndrome de Zellweger, y hemocromatosis perinatal, también conocida como hemocromatosis neonatal o enfermedad por almacenamiento de hierro neonatal.70 El secuestro de hierro en otros trastornos raros produce varios patrones de depósito de hierro localizado en la hemosiderosis pulmonar idiopática, la hemosiderosis renal y la enfermedad de Hallervorden-Spatz.71
Deficiencia de hierro
La deficiencia de hierro se refiere a condiciones en las que el hierro corporal está disminuido debido a un aumento sostenido en los requerimientos de hierro en comparación con su aporte. Los requerimientos de hierro incluyen no solo el hierro para restablecer las pérdidas fisiológicas y satisfacer las necesidades durante el crecimiento y el embarazo, pero tambin el que suple las pérdidas patológicas. La pérdida diaria fisiológica de hierro en el varón es un poco menor de 1.0 mg/día, y en la mujer normal que menstrúa de 1.5 mg/día.7 Pueden identificarse estadios secuenciales en la reducción del hierro corporal [ver figura 6 y tabla 1]. La reducción en las reservas de hierro sin un cambio en la cantidad de compuestos de hierro funcionales se denomina como reducción en la reserva de hierro. Cuando las reservas se agotan, se dice que los pacientes tienen depleción de hierro. La mayor disminución en el hierro corporal causa menor producción de hemoglobina y de otros compuestos funcionales que contienen hierro, estadio denominado eritropoyesis deficiente en hierro. La reducción aún mayor en el hierro corporal produce anemia por deficiencia de hierro. En todo el mundo, la deficiencia de hierro es la causa más común de anemia.39 Casi la tercera parte de la población mundial de alrededor de cinco billones tiene anemia, y la anemia por deficiencia de hierro causa o contribuye a la anemia en por lo menos 500 a 600 millones de personas.72
MANIFESTACIONES CLINICAS DE LA DEFICIENCIA DE HIERRO
Desde el punto de vista clínico los pacientes con deficiencia de hierro pueden estar asintomáticos y detectarse solo por resultados anormales en las pruebas de laboratorio. Otros pacientes pueden solicitar atención médica por las manifestaciones del trastorno subyacente que causó la deficiencia de hierro, pero pueden no tener datos relacionados con esta última. Un tercer grupo de enfermos puede sufrir signos y síntomas comunes a todas las anemias, como debilidad, mareo, fatiga fácil, palidez, irritabilidad y otras molestias indefinidas e inespecíficas. Por último, algunos pacientes presentan uno o más de los pocos signos y síntomas que se piensa son muy específicos de la deficiencia de hierro (escleras azules, coiloniquia y pagofagia).1 La deficiencia de hierro puede asociarse también con signos y síntomas que no se relacionan con anemia,73 como estomatitis angular, glositis, estenosis o pliegues esofágicos poscricoideos y atrofia gástrica. El síndrome de Plummer-Vinson, también conocido como síndrome de Paterson-Kelly,7 se manifiesta por glositis, ardor o sensación quemante en la boca, disfagia y deficiencia de hierro. La pagofagia, o pica con hielo, parece ser un síntoma muy específico de deficiencia de hierro y desaparece poco después de que se inicia el tratamiento.74 Otros tipos de pica pueden acompañar a la deficiencia de hierro, pero ninguno es tan específico como la pagofagia. En la coiloniquia, la porción distal de las uñas de los dedos adoptan forma de cuchara, y las uñas son delgadas y brillantes, quizá como consecuencia de trastornos en el crecimiento del lecho epitelial ungueal. Aunque se piensa que la coiloniquia es muy específica de la deficiencia de hierro, este signo ocurre en un número muy pequeño de pacientes. Las escleras con coloración azul se han descrito como un indicador muy sensible y específico de deficiencia de hierro.75 Esta coloración azulosa parece deberse a adelgazamiento de la esclerótica, quizá como consecuencia de una menor síntesis de colágena por la deficiencia de hierro. La deficiencia de hierro puede producir otras manifestaciones clínicas que no se relacionan con la anemia y que parecen ser causadas por la depleción de compuestos funcionales de hierro en los tejidos no eritroides. Las consecuencias no hematológicas de la falta de hierro incluyen menor tolerancia al ejercicio y desempeño laboral y deterioro en la inmunidad y resistencia a la infección. En los niños la deficiencia de hierro afecta en forma adversa el crecimiento, el desarrollo motor y las funciones cognoscitivas y de comportamiento. Estas alteraciones pueden no ser reversibles con el tratamiento posterior.39,73,76-78
CAUSAS DE DEFICIENCIA DE HIERRO
La pérdida de sangre es la causa más común de aumento en el requerimiento de hierro que origina deficiencia del mismo [ver tabla 3]. En los hombres y las mujeres posmenopáusicas, la deficiencia de hierro casi siempre es resultado de pérdida gastrointestinal.39 En las mujeres menstruando la pérdida genitourinaria suele ser la causa del aumento en los requerimientos de hierro. Los anticonceptivos orales pueden disminuir la pérdida menstrual de sangre, pero los dispositivos intrauterinos la aumentan. Deben considerarse también otras causas de hemorragia genitourinaria o respiratoria [ver tabla 3]. Para los donadores de sangre, cada donación significa una prdida de 200 a 250 mg de hierro. Durante los periodos de crecimiento en la infancia, niñez y adolescencia, los requerimientos de hierro pueden superar el aporte a partir de las dietas y reservas.73 El embarazo aumenta las necesidades de hierro y, sin hierro suplementario, causa una pérdida neta equivalente a alrededor de 1,200 a 1,500 ml de sangre. Después del parto, si el niño es alimentado al seno materno, la lactación aumenta los requerimientos de hierro en alrededor de 0.5 a 1.0 mg por día.
El aporte insuficiente de hierro puede contribuir al desarrollo de deficiencia de hierro. En los niños y mujeres con requerimientos altos de hierro, las dietas que contienen cantidades inadecuadas de hierro biodisponible pueden aumentar el riesgo de deficiencia. En los niños mayores, varones y mujeres posmenopáusicas, el aporte escaso de hierro en la dieta casi nunca es el único factor responsable de la deficiencia, por lo que deben buscarse otros factores etiológicos, en especial pérdida de sangre.
La menor absorción de hierro es una causa poco común de deficiencia. En algunos pacientes la malabsorción intestinal de hierro es solo un aspecto más de la malabsorción generalizada [ver tabla 3]. La cirugía gástrica, en especial la resección gástrica parcial o total o la gastroenterostomía para derivación del duodeno, puede causar deficiencia de hierro. Aunque la absorción de hierro en la dieta puede ser pobre, las sales terapéuticas se absorben bien, y la deficiencia puede corregirse con facilidad.
El riesgo de deficiencia de hierro es especialmente alto cuando los requerimientos aumentan y el aporte es inadecuado. Por ejemplo, los infantes alimentados con leche de vaca suelen tener deficiencia de hierro por la combinación de mayor pérdida por hemorragia gastrointestinal inducida por la leche de vaca y por las pequeñas cantidades de hierro biodisponible que existen en ésta.73 Las mujeres con requerimientos elevados de hierro por la menstruación suelen consumir dietas que tienen poco hierro biodisponible y con inhibidores de la absorción de hierro, como el calcio.
DIAGNOSTICO DE LA DEFICIENCIA DE HIERRO
La anemia por deficiencia de hierro es la única anemia microcítica hipocrómica asociada con ausencia en las reservas de hierro. En las otras anemias microcíticas hipocrómicas las reservas de hierro en la médula son normales o están aumentadas. Pueden usarse medidas indirectas del hierro corporal para identificar una secuencia característica de cambios que ocurren al disminuir el hierro corporal desde los niveles de reserva normal hasta los encontrados en la anemia por deficiencia de hierro [ver figura 6 y tabla 1]. La concentración de ferritina en plasma es la prueba más útil para la detección de la deficiencia de hierro porque las concentraciones de ferritina en plasma disminuyen al reducirse las reservas corporales.4 Una concentración de ferritina en plasma por debajo de 12 µg/L es virtualmente diagnóstica de ausencia de reservas de hierro. El hipotiroidismo y la deficiencia de ácido ascórbico pueden disminuir la concentración de ferritina en plasma en forma independiente de la reducción en el hierro corporal, pero estos trastornos no suelen constituir un problema diagnóstico.79 Por el contrario, la concentración normal de ferritina en plasma no confirma la presencia de reservas de hierro porque esta proteína puede aumentar en forma independiente por infecciones, inflamación, enfermedad hepática, neoplasias y otros trastornos. En estos casos la medición de la concentración del receptor de transferrina en plasma proporciona una manera de distinguir entre la anemia por deficiencia de hierro y la anemia asociada con trastornos inflamatorios crónicos.38 A diferencia de la concentración plasmática de ferritina, la concentración de receptor de transferrina en plasma no parece afectarse por infecciones, inflamación, enfermedad hepática o neoplasias, y puede ser un medio más eficaz para detectar anemia por deficiencia de hierro. La saturación de transferrina en plasma está disminuida tanto en la deficiencia de hierro como en los estados infecciosos e inflamatorios, y es de poca ayuda práctica para distinguir entre estos trastornos. Aunque rara vez se realiza examen de la médula ósea solo para evaluar el estado del hierro, casi siempre puede verificarse el estado de deficiencia de hierro por la evaluación directa de las reservas de este elemento en la médula. Si no existen reservas de hierro se establece diagnóstico de deficiencia de hierro, si se encuentra hemosiderina, se excluye esta posibilidad. Además, en la deficiencia de hierro no existen sideroblastos en la médula, o estos están muy disminuidos, constituyendo menos del 10 porciento de los normoblastos. Una prueba terapéutica con hierro puede ser un medio eficaz de establecer el diagnóstico de deficiencia de hierro.
TRATAMIENTO DE LA DEFICIENCIA DE HIERRO
El tratamiento para la anemia por deficiencia de hierro consiste en corregir tanto la deficiencia de hemoglobina como reemplazar las reservas de hierro.39,73 Para casi todos los pacientes el hierro oral es el tratamiento de elección. El tratamiento con hierro oral es eficaz, seguro y económico. Debido al riesgo de reacciones adversas locales y sistémicas, el hierro parenteral solo debe usarse en el pequeño número de pacientes que no pueden absorber o tolerar el hierro oral o que tienen requerimientos de hierro que no pueden satisfacerse por el tratamiento oral debido a hemorragia crónica no controlable. En la anemia severa por deficiencia de hierro puede requerirse transfusión de eritrocitos para los pacientes en los que la velocidad de pérdida crónica de hierro excede la velocidad a la que puede reemplazarse el hierro con tratamiento parenteral. Aunque la mayoría de los pacientes toman el hierro oral sin dificultad, el 10 a 20 porciento sufre efectos colaterales, principalmente de tipo gastrointestinal.
La deficiencia de hierro casi siempre puede tratarse en forma eficaz, y el tratamiento oral y parenteral da resultados semejantes [ver tabla 4]. Ocurre mejoría de los síntomas en los primeros días de tratamiento. En la anemia no complicada, la respuesta hematológica inicial, que consiste en reticulocitosis leve, comienza en tres a cinco días después del inicio del tratamiento, alcanza un máximo a los ocho a 10 días y disminuye después. Después de la primera semana la concentración de hemoglobina comienza a aumentar y suele ser normal alrededor de seis semanas después. La microcitosis puede no resolverse por completo hasta después de cuatro meses. Si la deficiencia de hierro se trata con hierro oral en dosis de 200 mg/día o menos, la concentración de ferritina en plasma permanece en menos de 12 µg/L hasta que la anemia se corrija y después aumenta en forma gradual al repletarse las reservas de hierro. Si la respuesta al tratamiento con hierro no es completa y característica, estará indicado reevaluar al paciente. Uno de los problemas más comunes es confundir la anemia de una enfermedad crónica con anemia por deficiencia de hierro. La recuperación puede ser más lenta cuando coexisten padecimientos, incluyendo otras deficiencias nutricionales, enfermedad hepática o renal, infecciones, inflamación o neoplasias, o pérdida de sangre oculta continua. Si el paciente fue tratado con hierro oral, deberán revisarse la forma y dosis de hierro, la constancia al tratamiento y la posibilidad de malabsorción.
DR. GARY M. BRITTENHAM
Función del eritrocito
La función principal del eritrocito es el transporte de oxígeno de los pulmones a los tejidos periféricos para su utilización y del bióxido de carbono a los pulmones para su excreción.1,2 El eritrocito maduro dedica más del 95 porciento de su proteína intracelular, la hemoglobina, a esta tarea. La hemoglobina, que es la molécula transportadora del oxígeno, se une a las moléculas de oxígeno en el ambiente de alta tensión de oxígeno de los alveolos pulmonares y libera moléculas de oxígeno en los tejidos periféricos en donde la tensión de oxígeno es baja. En el eritrocito, se encuentra dentro de una bicapa lípida sostenida por un citoesqueleto que determina la forma bicóncava y discoide de esta célula. La forma del eritrocito optimiza el paso de la célula a través del sistema circulatorio y permite la aposición de los eritrocitos y las células parenquimatosas a través del delgado endotelio de los capilares, lo que facilita el intercambio de oxígeno y bióxido de carbono. Sin eritrocitos, la sangre del plasma transportaría solo alrededor de 5 ml de O2/L, pero con una hemoglobina normal la sangre total puede transportar alrededor de 200 ml de O2/L.2
ESTRUCTURA DE LA HEMOGLOBINA
La hemoglobina, que tiene un peso molecular de 64,400 daltons, es un tetrámero compuesto de dos pares de cadenas polipeptídicas globinas diferentes con un grupo hem, la ferroprotoporfirina IX, unido en forma covalente a un sitio específico en cada cadena.3 La principal hemoglobina adulta, la hemoglobina A, está formada por un par de cadenas a (cada una conteniendo 141 aminoácidos) y un par de cadenas b (cada una con 146 aminoácidos), y se expresa como a2b2. Su estructura tridimensional es esferoidal, con un solo eje de asimetría y los cuatro grupos hem en surcos colocados equidistantes alrededor de la superficie de la molécula.4 La configuración deoxi de la hemoglobina se estabiliza por uniones intra e intercadenas, algunas de estas dependen de la unión de protones y otras de la presencia de 2,3-difosfoglicerato (2,3-DPG). Estas uniones se rompen en forma secuencial durante la oxigenación. Con la unión de una molécula de oxígeno a un grupo hem cada subunidad a o b no unida sufre un cambio en su estructura terciaria, lo que produce un cambio en la configuración de toda la molécula de hemoglobina [ver figura 1]. Estos cambios de conformación producen un aumento en la afinidad por oxígeno del resto de las subunidades no unidas, un fenómeno denominado cooperatividad o interacción hem-hem.5,6 Por el contrario, si el oxígeno es liberado de un hem en la oxihemoglobina, el resto de los sitios unidos sufren cambios que disminuyen su afinidad por el oxígeno, facilitando la liberación de esta molécula. Estos cambios conformacionales son también los responsables de la menor afinidad de la hemoglobina por el oxígeno al disminuir el pH, el llamado efecto Bohr.3 La oxigenación disminuye los valores K de los grupos ácidos en residuos específicos de aminoácidos, liberando alrededor de 2.8 protones por molécula de hemoglobina. El efecto Bohr es fisiológicamente benéfico tanto en los pulmones, en donde la liberación de bióxido de carbono aumenta el pH, aumentando la afinidad por oxígeno y su captación, como en los tejidos, en donde la captación del bióxido de carbono disminuye el pH, reduciendo la afinidad por oxígeno y facilitando su liberación.
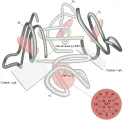
|
| Figura 1 |
| Molécula de hemoglobina |
CURVA DE DISOCIACION DE OXIGENO-HEMOGLOBINA
La curva de disociación de oxígeno-hemoglobina [ver figura 2] es una gráfica del equilibrio entre el oxígeno y la hemoglobina a varias tensiones de oxígeno (PO2). La forma sigmoidea de la curva, que es resultado de la interacción entre las subunidades de la hemoglobina humana, tiene ventajas fisiológicas.1-3 En los pulmones, con una presión parcial de oxígeno de alrededor de 90 mm Hg, la hemoglobina está saturada al 97 porciento. Después de dejar el oxígeno en los tejidos, a una PO2 de alrededor de 40 mm Hg en la sangre venosa mixta, la saturación de la hemoglobina es de 75 porciento. Por lo tanto, se libera una cantidad relativamente grande de oxígeno con una disminución discreta en la tensión de oxígeno. La P50, que es la presión parcial de oxígeno a la que la hemoglobina está saturada a la mitad, es una medida útil de la afinidad de la hemoglobina por el oxígeno: a mayor afinidad menor P50. En condiciones fisiológicas (i.e., temperatura de 37C [98.6F], pH de 7.40, 2,3-DPG de 5 mmol/L y tensión de bióxido de carbono [PCO2] de 40 mm Hg), la P50 de la sangre del adulto normal es de 26±1 mm Hg. La P50 disminuye (se desplaza a la izquierda) con el aumento en el pH, la reducción en el 2,3-DPG o la reducción en la temperatura.

|
| Figura 2 |
| Curva de disociación de oxígeno-hemoglobina |
EFECTOS DEL 2,3-DIFOSFOGLICERATO
El producto intermedio de la glucólisis, 2,3-DPG, un anión con carga importante, está presente en los eritrocitos maduros en una concentración de 4 a 5 mmol/L, aproximadamente la misma concentración celular que la hemoglobina. Una molécula de 2,3-DPG se une a una molécula de desoxihemoglobina en el hueco formado por los extremos amino de las cadenas b, reduciendo la afinidad del oxígeno por la hemoglobina al estabilizar la conformación desoxi1-3 sin alterar la interacción o el efecto Bohr. El trifosfato de adenosina (ATP) se une a la hemoglobina en forma semejante, pero la mayoría del ATP intracelular forma un complejo con Mg2+, que bloquea la reacción con la desoxihemoglobina. Como resultado, el 2,3-DPG es el regulador alostérico cuantitativamente más importante de la afinidad por el oxígeno.2 En presencia de hipoxia (por anemia aguda, enfermedad cardiopulmonar, otros trastornos o exposición súbita a altitudes elevadas), la concentración de 2,3-DPG aumenta en horas, incrementando la P50. El desplazamiento de la curva de disociación de oxígeno-hemoglobina a la derecha facilita la entrega de oxígeno a los tejidos, pero a costa de disminuir su adquisición en los pulmones. Los aumentos en el 2,3-DPG pueden ayudar a adaptarse a corto plazo al estrés hipóxico si el aporte de oxígeno es abundante y la reserva cardiopulmonar robusta. A altitudes elevadas, cuando el sistema cardiovascular es incapaz de satisfacer en forma eficaz la mayor demanda circulatoria, o en otras circunstancias patológicas, el aumento en la cantidad de 2,3-DPG puede ser contraproducente.
APORTE DE OXIGENO A LOS TEJIDOS
El eritrocito circulante es un elemento crucial en el transporte de oxígeno a los tejidos, pero una variedad de otros componentes fisiológicos deben funcionar también en forma integrada para proporcionar un aporte adecuado de oxígeno.2 El volumen y la viscosidad de la sangre afectan el aporte de oxígeno. La concentración de eritrocitos circulantes depende, en parte, de la actividad eritropoyetica, que a su vez es regulada por la producción renal de eritropoyetina. Otros componentes importantes incluyen la tensión de oxígeno en el ambiente, los factores pulmonares que influyen en la oxigenación y la ventilación, factores cardiovasculares como el gasto cardiaco y el flujo sanguíneo regional y la magnitud de oxígeno que no se libera en los tejidos.
TRANSPORTE DE BIOXIDO DE CARBONO
La hemoglobina se une al bióxido de carbono después de dejar oxígeno en los tejidos. La mayoría del bióxido de carbono de los capilares tisulares es transportado a los pulmones como bicarbonato, y alrededor de un 10 porciento es transportado como un complejo carbamino unido en forma reversible a los grupos aminoterminales de las cadenas de globina. La desoxihemoglobina tiene mayor afinidad por el bióxido de carbono que la oxihemoglobina, lo que facilita su transporte desde los tejidos para eliminarse en el pulmón.
Trastornos del metabolismo del hierro
Todas las células del organismo requieren hierro. En el metabolismo humano el hierro se usa para transportar y almacenar oxígeno, transportar electrones y actuar como el elemento catalítico cuantitativamente más importante, con funciones cruciales en el metabolismo oxidativo y en el crecimiento y proliferación celular. En los humanos el hierro se encuentra casi en forma exclusiva como ión ferroso (Fe2+) o férrico (Fe3+) y puede cambiar con facilidad entre estos dos estados de oxidación. Debido a que estas formas del hierro son muy reactivas, casi siempre existe como un componente de complejos hierro-porfirina o metaloenzimas, o unido a proteínas especializadas de transporte o almacén. Estos compuestos evitan o limitan la aparición de hierro en forma no unida o inorgánica, evitando la participación del hierro en reacciones de producción de radicales libres que pueden lesionar los componentes celulares. Bajo condiciones fisiológicas, la cantidad de hierro en el organismo está controlada dentro de límites estrechos para evitar o su deficiencia o su exceso. Cuando existe deficiencia de hierro el organismo es incapaz de sintetizar suficiente cantidad de hem, otros complejos hierro-porfirina, metaloenzimas u otros compuestos que contienen hierro para realizar las funciones normales. Con la sobrecarga de hierro, el exceso puede provocar reacciones dañinas que causan daño progresivo y eventualmente letal a órganos vitales.
METABOLISMO DEL HIERRO
Normalmente la concentración de hierro en el organismo es de 40 mg Fe/kg de peso corporal en la mujer y alrededor de 50 mg Fe/kg en el hombre. La mayoría de este hierro es un componente de varios compuestos funcionales dedicados al transporte, entrega, utilización o consumo de oxígeno. La mayor proporción de hierro se encuentra en el eritrón como hierro de la hemoglobina (28 mg/kg en mujeres y 32 mg/kg en hombres), dedicado al transporte y entrega de oxígeno. Se encuentran también cantidades pequeñas de hierro eritrón (<1mg/kg) en el hem y en enzimas no hem en eritrocitos en desarrollo.
El resto del hierro funcional se encuentra como hierro de mioglobina (4 mg/kg en mujeres y 5 mg/kg en hombres) en el músculo y en enzimas que contienen o dependen de hierro (1 a 2 mg/kg) en todas las células del organismo. Se depositan pequeñas cantidades de hierro dentro de ferritina en las células eritroides, pero el principal almacen de hierro (5 a 6 mg/kg en mujeres y 10 a 12 mg/kg en hombres) tiene lugar en hepatocitos y macrófagos del hígado, médula ósea, bazo y músculo [ver figura 3]. La pequeña cantidad de hierro en transporte (alrededor de 0.2 mg/kg) en el plasma y el líquido extracelular está unido a la proteína transferrina, que lleva el hierro para satisfacer las necesidades tisulares en todo el organismo.7,8
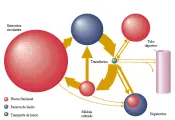
|
| Figura 3 |
| Aporte y almacén del hierro corporal en el organismo |
El eritrón se define como la totalidad de elementos eritroides en todas las fases de desarrollo y los tejidos en los que se producen.1 La mayoría del hierro en el organismo se localiza en el eritrón, y la mayoría del flujo diario de hierro pasa a través de este compartimiento. La vía predominante de flujo interno de hierro es unidireccional de la transferrina del plasma al eritrón, al sistema monocito-macrófago y después de regreso a la transferrina.
El flujo de hierro hacia y desde el eritrón constituye alrededor de las cuatro quintas partes del hierro que pasa a través del compartimiento de transferrina cada día. El reciclamiento eficaz del hierro de los eritrocitos recién destruidos o formados es una característica distintiva del metabolismo humano del hierro.
En condiciones fisológicas, menos del 0.05 porciento del hierro corporal total se adquiere o pierde cada día. La eficacia con la que el hierro se recicla en el organismo es única, ningún otro animal conserva el hierro en forma tan eficaz. Fisiológicamente, el eritrón adquiere hierro de transferrina a través de un receptor específico de transferrina localizado en la superficie de la membrana de las células eritroides inmaduras. La mayoría de este hierro se usa para la síntesis de hemoglobina y después entra a la circulación en los eritrocitos. Pequeñas cantidades de hierro se almacenan en la ferritina (la proteína almacenadora de hierro), entran a enzimas que contienen hierro de las células eritroides inmaduras o se pierden en los productos de la eritropoyesis ineficaz.
Los eritrocitos viejos son fagocitados por macrófagos especializados en el bazo, la médula ósea y el hque regresan la mayoría del hierro eritroide catabolizado al compartimiento de transferrina, donde comienza su ciclo de nuevo. La fagocitosis de las células eritroides inmaduras y de los eritrocitos viejos explica la mayoría de todo el hierro almacenado que se encuentra en los macrófagos del hígado, médula ósea y bazo, ya que nada o casi nada de este hierro deriva de la transferrina.
Por el contrario, las células parenquimatosas del hígado pueden captar hierro de la transferrina o entregarlo a la misma. La quinta parte restante del hierro que se moviliza hacia y desde la transferrina corresponde a cantidades más o menos iguales de hierro absorbido y perdido, que se desplaza de los compartimientos de transferrina del plasma y extravascular, se mueve hacia y desde los hepatocitos y se intercambia entre la transferrina extravascular y los tejidos parenquimatosos.
BALANCE DEL HIERRO
El balance del hierro es el resultado de la diferencia entre la cantidad de hierro que recibe y que pierde el organismo. Debido a que los humanos son incapaces de excretar el exceso de hierro, el balance está regulado fisiológicamente por el control de la absorción.9 Los dos principales factores que influyen en la absorción del hierro son el nivel de las reservas corporales de hierro y la magnitud de la eritropoyesis. Si la reserva de hierro aumenta la absorción disminuye y viceversa. La absorción aumenta al aumentar la actividad eritropoyética, en especial con la eritropoyesis ineficaz. El hierro de la dieta se absorbe principalmente por las células mucosas del duodeno y la porción superior del yeyuno. La base molecular para la absorción de hierro por las células mucosas de la porción superior del intestino delgado sigue siendo incierta, pero existen evidencias de un mecanismo mucina-mobilferrina-integrina.10
TRANSPORTE, ALMACEN Y REGULACION DEL HIERRO
Desde el punto de vista fisiológico, tres proteínas (transferrina, receptor de transferrina y ferritina) son responsables del transporte y almacén del hierro. A su vez, la expresión coordinada de estas tres proteínas está controlada por una cuarta clase de proteínas, las proteínas reguladoras del hierro (PRH). La transferrina es la proteína transportadora que proporciona el único medio fisiológico para el transporte de este elemento a las células. El receptor de transferrina es el receptor de la superficie celular que permite la entrada del complejo transferrina-hierro a las células. La ferritina, la proteína de almacén, proporciona un reservorio en la célula en el que puede depositarse el hierro hasta que se requiera para el metabolismo. Dentro de las células, la expresión del receptor de transferrina y de la ferritina está regulado por un ácido ribonucleico mensajero (ARNm) de proteínas de unión identificadas actualmente como proteínas reguladoras del hierro. Cada una de las proteínas principales ha sido aislada, purificada y caracterizada [ver figura 4].
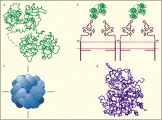
|
| Figura 4 |
| Proteínas reguladoras del hierro |
TRANSFERRINA
La transferrina [ver figura 4] transporta el hierro a los eritrocitos en desarrollo y a otras células del organismo, en especial las que están en crecimiento o proliferación.11,12 En los humanos el hepatocito es la fuente de casi toda la apotransferrina circulante, que se divide en porciones más o menos iguales entre el plasma y los líquidos extravasculares.
La apotransferrina es una glucoproteína transportadora con vida media de alrededor de ocho días,11 después de entregar el hierro a las células, la apoferritina retorna con rapidez al plasma para funcionar de nuevo como transportadora. La apotransferrina es una glucoproteína de una sola cadena bilobular con dos lóbulos semejantes en los extremos N- y C-terminales.13 Debido a que un solo ión férrico puede unirse en cada lóbulo, existen cuatro formas de la molécula: (1) apotransferrina, (2) transferrina monomérica, con un átomo de hierro unido al lóbulo N-terminal (FeNTf), (3) transferrina monoférrica, con un átomo de hierro unido al lóbulo C-terminal (FecTf), y (4) transferrina diférrica (Fe2Tf). La saturación de la transferrina es la proporción de sitios disponibles para unión de hierro en la transferrina que están ocupados por átomos de hierro, expresado en porcentaje. Las proporciones relativas de apotransferrina, transferrina monoférrica y transferrina diférrica son una función estocástica de la saturación de transferrina.
RECEPTOR DE TRANSFERRINA
El receptor de transferrina [ver figura 4] es la única vía de entrada fisiológica a la célula para el hierro unido a transferrina, y también ayuda a liberar el hierro de la transferrina para su uso en la célula.14,15 Los receptores de transferrina se localizan en la superficie de la membrana de todas las células nucleadas, en números que dependen de los requerimientos celulares de hierro. Para cada célula determinada, el determinante predominante del aporte de hierro a la célula es el número de receptores de transferrina expresados en la superficie de la célula. Por ello, el número de receptores de transferrina es más alto en las células de la médula eritroide, el hígado y la placenta. Desde el punto de vista estructural, el receptor de transferrina consiste en dos subunidades transmembrana glucoproteicas idénticas unidas por un puente disulfuro [ver figura 4]. La eficacia con la que el receptor de transferrina puede entregar el hierro unido a la transferrina depende del contenido de hierro de la transferrina. Cada subunidad puede fijar una molécula de transferrina, de modo que el receptor puede aceptar dos moléculas. El receptor dimérico puede entonces permitir la entrada de cuatro átomos de hierro si cada transferrina es diférrica o solo dos átomos de hierro si cada una es monoférrica. Tanto el pH como el contenido de transferrina determinan la magnitud de la afinidad del receptor de transferrina por la transferrina. A un pH fisiológico de 7.4, con suficiente transferrina con hierro para saturar los receptores disponibles, el receptor de transferrina casi no tiene afinidad por la apotransferrina, afinidad moderada por la transferrina monoférrica y la mayor afinidad por la transferrina diférrica (más de cuatro veces mayor que para la transferrina monoférrica).11 Al disminuirse el pH a alrededor de 5, la afinidad del receptor de transferrina por la apotransferrina aumenta a una magnitud equiparable a la que tiene por la transferrina diférrica a un pH de 7.4.
FERRITINA
La ferritina [ver figura 4] es la principal proteína para almacén de hierro y tiene una vida media de alrededor de 60 horas (para ferritina hepática). Casi todas las células contienen ferritina, que funciona tanto como un sitio de almacén seguro para hierro como una reserva accesible que ha sido adquirida por la célula ante la existencia en exceso a sus necesidades metabólicas inmediatas. En consecuencia, las mayores cantidades de ferritina se encuentran en las células dedicadas al almacén de hierro (i.e., macrófagos y hepatocitos) y en las células con los mayores requerimientos para la síntesis de compuestos que contienen hierro (i.e., células eritroides en desarrollo).16 La apoferritina, que tiene un peso molecular de 440,000, es una concha esférica con radio de alrededor de 6 mm, cada molécula puede almacenar hasta 4,500 átomos de hierro en el interior en un centro polinuclear de hidrofosfato férrico. La proteína de apoferritina está formada por 24 subunidades oblongas que se denominan como H (pesadas, con un peso molecular de 21,100 daltons) y L (o ligeras, con un peso molecular de 19,700 daltons). Las subunidades tienen homología considerable, pero existe un sitio de ceruloplasmina en la subunidad H17 y no en la subunidad L. La abundancia relativa de subunidades H y L está determinada tanto por el tipo celular específico como por el estado férrico de la célula. El mayor número de subunidades L se encuentra en las células de tejidos que tienen función especializada de almacén de hierro, como el hígado y el bazo,18 mientras que existen más subunidades H en células de tejidos que no funcionan como depósitos de hierro, como el músculo cardiaco. Para un tipo específico de célula, se encuentra mayor proporción de subunidades L cuando es mayor el almacén de hierro. Estas observaciones sugieren que las ferritinas con mayor proporción de subunidades H son más activas en el metabolismo y que las que tienen mayor abundancia de subunidades L sirven para almacenar el hierro a más largo plazo.
PROTEINAS REGULADORAS DE HIERRO
Las proteínas críticas que permiten al hierro autorregular su disponibilidad intracelular son las proteínas reguladoras del hierro PRH1 y PRH2 [ver figura 4]. Las proteínas reguladoras del hierro son proteínas que unen ARN citoplásmico que funcionan en una forma de acción-trans sobre los ARNm que contienen un elemento que responde al hierro (ERH), una estructura reguladora de acción-cis.19,20 El ERH, que es el sitio de unión para las PRH, es una estructura en forma de pasador compuesta de una secuencia conservada de 28 nucleótidos. La región 3í no traducida del ARNm del receptor de transferrina, la región 5í no traducida del ARNm de la ferritina y el ARNm de la forma eritroide específica de la sintetasa de ácido d-aminolevulínico (AAL) contienen ERH.20 Los tejidos que contienen PRH muestran diferencias importantes en la cantidad de PRH1 y PRH2. Los tejidos con la mayor cantidad de almacén de hierro, como el hígado y el bazo, tienen mayor cantidad de PRH1.21 Los tejidos con mayor utilización del hierro, como el corazón y el músculo esquelético, contienen la mayor cantidad de la PRH2.22
Se ha encontrado que la PRH1 es la aconitasa citosólica23 (hidro-liasa de citrato [isocitrato]), una proteína con sulfuro de hierro que actúa enzimáticamente dentro del ciclo de Krebs en la interconversión de citrato a isocitrato. Dentro de la clula, la aconitasa existe en una forma citoplásmica (codificada por un gen en el cromosoma 9) y mitocondrial (codificada por un gen en el cromosoma 22). Tanto las formas citoplásmica como mitocondrial son enzimáticamente activas como enzima aconitasa, pero solo la forma citoplásmica puede funcionar como PRH. Un sitio específico (4Fe-4S) forma el sitio catalíticamente activo de la enzima aconitasa, en el que tres átomos de hierro se unen a la estructura de la proteína y el cuarto átomo se une a un azufre inorgánico en el sitio específico. Los cambios en la disponibilidad de hierro intracelular parecen alterar la conformación del sitio Fe-S, transformando de modo reversible la aconitasa enzimáticamente activa, que normalmente es incapaz de unirse al ARN con gran afinidad, en una forma que tiene gran afinidad de unión al ARN y que no tiene actividad de aconitasa. Estas conversiones permiten a la PRH1 monitorizar los cambios en la disponibilidad del hierro intracelular. La mayor cantidad de hierro intracelular aumenta las cantidades de PRH1 en la forma de baja afinidad de unión al ARN, disminuyendo la unión de la PRH1 al ERH. La reducción en el hierro intracelular disponible aumenta la cantidad de PRH1 en la forma de alta afinidad de unión al ARN, aumentando la unión de la PRH1 al ERH.
La PRH2 no tiene actividad de aconitasa, pero parece unirse al ERH con una afinidad indistinguible de la de la PRH1 y funcionar con la misma eficacia como represor de la traducción in vitro.22 La reducción en la disponibilidad de hierro intracelular aumenta la actividad de unión al ARNm de ambas PRH, pero por diferentes motivos.24 El control de la PRH1 es principalmente postraducción, y el cambio de la forma de no unión (con actividad de aconitasa) a la forma de unión al PRH (sin actividad de aconitasa) produce poco cambio en la cantidad de PRH1. Por el contrario, la mayor actividad de unión al ARNm de la PRH2 es el resultado de síntesis de novo.25 Las consecuencias de las diferencias en la distribución tisular y en los mecanismos de regulación del hierro de las PRH1 y PRH2 aún no se conocen, pero la dos PRH parecen funcionar de modo semejante como sensores de la disponibilidad de hierro intracelular.26
APORTE Y ALMACEN DE HIERRO EN LA CELULA
La transferrina, el receptor de transferrina y la ferritina actúan juntos para proporcionar hierro a la célula [ver figura 5].11 Al inicio dos moléculas de transferrina (monoférrica o diférrica) se unen a un receptor de transferrina sobre la superficie celular. Con la eritropoyesis normal, la mayoría del aporte de hierro al eritrón es proporcionado por transferrina diférrica cuando la saturación de transferrina es de 33 porciento o mayor. Con saturaciones de transferrina de 19 porciento o menos la mayoría del hierro proviene de transferrina monoférrica. Ambas formas de transferrina aportan hierro a una saturación de 20 a 32 porciento. El complejo hierro-transferrina-receptor de transferrina se agrega a otros complejos en un orificio cubierto por clatrin?, que después es translocado al interior de la célula como una vesícula cubierta. Después de que el clatrin? es separado, la vesícula sin cubierta se fusionan con otras vesículas para formar un endosoma multivesicular. El endosoma se acidifica hasta un pH de 5.6 por una bomba de protones mientras viaja en el interior de la célula. Con la caída en el pH, el receptor de transferrina sufre cambios de conformación que ayudan a liberar el hierro de la transferrina mono o diférrica. Aunque aún no se conocen los mecanismos moleculares que participan, el hierro disociado es entonces transportado a través de la membrana endosómica para ser utilizado en la síntesis de compuestos que contengan hierro o para su almacén en la ferritina citoplásmica. En el endosoma a un pH ácido, la apotransferrina sin hierro se une en forma ávida al receptor de transferrina, y el complejo es transportado de regreso a la membrana celular en el endosoma. Aunque algunos endosomas se desvían hacia el aparato de Golgi para su resialilación y otros son reparados, la mayoría de los endosomas llegan en forma directa a la superficie celular y se fusionan con la membrana. El complejo apotransferrina-receptor de transferrina se expone entonces al pH de 7.4 del plasma, lo que causa pérdida de la afinidad de la apotransferrina por el receptor. La apotransferrina entra al reservorio circulante en el plasma y el receptor de transferrina está de nuevo disponible para unir a una transferrina que lleve hierro.11
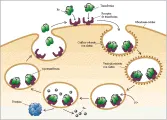
|
| Figura 5 |
| Mecanismos de aporte de hierro |
REGULACION DE LA CAPTACION DE HIERRO EN LA CELULA Y SU ALMACEN POR PROTEINAS REGULADORAS DE HIERRO
Las PRH controlan la captación intracelular y el almacén del hierro al controlar la traducción en la síntesis del receptor de transferrina y de la ferritina.19,20 Las PRH1 y PRH2 responden en forma semejante a cambios en el hierro disponible dentro de la célula y parecen fijarse al ERH con igual avidez y eficacia.26 La síntesis del receptor de transferrina se regula al controlar la estabilidad del ARNm citoplásmico del receptor de transferrina. Existen tres ERH en la región no traducida 3í (RNT3í) del ARNm. Al unirse la PRH al ERH en la RNT3í, la estabilidad del ARNm aumenta y se reduce su degradación citoplásmica. La cantidad de ARNm citoplásmico del receptor de transferrina aumenta, y la velocidad de síntesis del mismo también. Al incrementarse el número de receptores de transferrina en la superficie celular, la captación celular de hierro será mayor. La síntesis de ferritina esta regulada por un control en la traducción del ARNm de ferritina en el citoplasma. Existe un solo ERH en la región no traducida 5í (RNT5í) del ARNm de la ferritina. Cuando este ERH en la RNT5í se une a una PRH, se detiene la traducción del ARNm de la ferritina. Al producirse menos ferritina disminuye el almacén de hierrro y aumenta la disponibilidad de hierro celular. Por lo tanto, los cambios en la cantidad de la PRH tienen efectos opuestos en la producción del receptor de transferrina y la ferritina. Si cae el nivel de hierro intracelular, la cantidad de PRH de alta afinidad aumenta. A mayor cantidad de PRH la unión a los ERH en la RNT3í estabiliza al ARNm del receptor de transferrina, aumentando la producción del receptor de transferrina y la disponibilidad de hierro intracelular. El aumento en las PRH también disminuye la síntesis de ferritina al unirse a la RNT5í del ARNm , disminuyendo el almacén de hierro y aumentando la disponibilidad de hierro intracelular. Por el contrario, el aumento en el hierro intracelular disminuye la cantidad de PRH de alta afinidad, disminuyendo la captación de hierro por la célula al reducir la síntesis de la proteína del receptor de transferrina y aumenta el almacén de hierro al incrementar la síntesis de proteína ferritina. De esta forma coordinada, las PRH actúan para conservar la disponibilidad de hierro intracelular al controlar la traducción en la síntesis de las proteínas encargadas de la captación y almacenamiento del hierro.19,20
Evaluación de laboratorio del metabolismo del hierro
Se observan cambios característicos en la función, transporte y almacén de hierro al aumentar o disminuir su cantidad en el organismo [ver figura 6 y tabla 1]. Pueden usarse métodos tanto directos como indirectos para evaluar el estado corporal del hierro, pero ningún indicador o combinación de indicadores proporciona una forma óptima para evaluar al aporte y almacén del hierro en el organismo en todas las situaciones. Además, los métodos disponibles deben interpretarse tomando en cuenta como se afecta cada medida por la presencia concomitante de inflamación, infección, neoplasias, enfermedad hepática, deficiencia de ácido ascórbico o desnutrición.
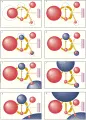
|
| Figura 6 |
| Reservas y distribución del hierro |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
* Puede ser normal en algunos pacientes |
MEDIDAS DIRECTAS DEL ESTADO DEL HIERRO CORPORAL
Las medidas directas del hierro corporal proporcionan la manera cuantitativa de evaluar a este elemento y tienen una gran sensibilidad y especificidad. La flebotomía cuantitativa permite calcular las reservar de hierro que pueden movilizarse para la producción de hemoglobina. Este método típicamente requiere la eliminación de alrededor de 500 ml de sangre una o dos veces por semana hasta que se agoten las reservas de hierro, lo que se indica por una reducción sostenida de la concentración de hemoglobina a menos de 10 g/dl sin que se continúen las flebotomías. El hierro almacenado y movilizado puede calcularse a partir de la cantidad de hierro de la hemoglobina extraído, corrigiendo por la diferencia entre las concentraciones inicial y final de hemoglobina y por la absorción de hierro de la dieta durante el periodo de flebotomía. La flebotomía cuantitativa puede ser útil para establecer el diagnóstico retrospectivo de hemocromatosis asociada al HLA en homocigotos y de otras formas de sobrecarga de hierro. Este método no puede usarse en la mayoría de las formas de anemia con exceso de hierro o en pacientes dependientes de transfusión, y no es útil en pacientes con reducción en la cantidad del hierro corporal. La aspiración y biopsia de médula ósea son de mayor utilidad en la evaluación de los pacientes con deficiencia de hierro u otras condiciones en las que se requiere evaluar el aporte de hierro para la eritropoyesis y las características morfológicas de la hematopoyesis. La tinción de la médula con azul de Prusia puede proporcionar información sobre (1) las reservas de hierro en los macrófagos al evaluar la cantidad de hemosiderina en la médula y (2) la morfología y proporción de sideroblastos en la médula (eritroblastos que contienen uno o mas gránulos positivos al azul de Prusia, que pueden identificarse por microscopía de luz). Si se requiere, puede medirse también el almacén de hierro en los macrófagos en una muestra de médula por medio de la determinación química del hierro no hem. El estudio de la médula ósea no proporciona datos sobre el grado de acúmulo de hierro en las células parenquimatosas, por lo que no suele ser útil en la evaluación de la sobrecarga de hierro. La biopsia hepática con determinación cuantitativa de hierro y estudio histoquímico es el mejor método para evaluar el hierro corporal en pacientes con todas las formas de sobrecarga sistémica de hierro.27 En este grupo de pacientes, la determinación cuantitativa de la concentración de hierro no hem en el tejido hepático obtenido por biopsia es el método de referencia para la medición del acúmulo de hierro en el organismo. El examen del patrón de depósito de hierro en los hepatocitos y las células de Kupffer en la muestra de biopsia proporciona información sobre la magnitud relativa del acúmulo de hierro en las células parenquimatosas y los macrófagos. Aunque estos métodos directos para evaluar el estado del hierro pueden proporcionar resultados definitivos en las circunstancias apropiadas, todos son métodos invasivos que pueden ser inaceptables, incómodos y, en el caso de la biopsia hepática, riesgosos para los pacientes. Se han examinado varias alternativas no invasivas, incluyendo la determinación de la susceptibilidad magnética hepática,28-30 la tomografía computada,31 y la imagen por resonancia magnética,32 pero en la actualidad estos procedimientos están en fase de investigación.
CALCULO INDIRECTO DEL ESTADO DEL HIERRO CORPORAL
Comparados con los métodos directos de medición del hierro corporal, los métodos indirectos son menos cuantitativos y no son tan sensibles ni tan específicos porque todos los métodos indirectos se afectan por diversas condiciones. A pesar de estas limitantes, los indicadores indirectos son accesibles y requieren solo una muestra de sangre u orina. La concentración de ferritina en plasma es la medición más empleada como indicador indirecto de las reservas corporales de hierro.33 En personas sanas, las concentraciones de ferritina en plasma suelen disminuir al disminuir la reserva de hierro y aumentan cuando éste se acumula [ver figura 6 y tabla 1]. Por el contrario, en personas con diversos padecimientos, la relación entre el hierro en plasma y el corporal puede estar muy alterado.34 Por ejemplo, la deficiencia de ácido ascórbico puede disminuir la concentración de ferritina en plasma sin que cambien las reservas corporales de hierro. En ausencia de deficiencia de ácido ascórbico, y quizá de hipotiroidismo y desnutrición severa, una concentración de ferritina en plasma de menos de 12 µg/L es muy específica de deficiencia de hierro. Las concentraciones de ferritina en plasma que son normales o aumentadas pueden ser difíciles de interpretar porque la ferritina plasmática es un reactante de fase aguda, lo que significa que su síntesis y concentración plasmática aumenta con la infección, la inflamación, la enfermedad hepática aguda y crónica y las neoplasias. En algunos casos estos trastornos también provocan liberación de ferritina de células dañadas (en especial con hepatopatía o quimioterapia citotóxica) o interfieren con la depuración de la ferritina circulante. Este aumento en la ferritina del plasma puede ocultar la deficiencia de hierro o sugerir en forma falsa exceso del mismo. Se ha sugerido que la concentración máxima de ferritina en plasma en respuesta a un aumento en el hierro corporal es de alrededor de 4,000 µg/L, lo que quizá representa el límite fisiológico superior de síntesis.35 Pueden presentarse concentraciones por arriba de 4,000 µg/L por la liberación de ferritina intracelular de células dañadas. La ferritina plasmática puede aumentar también por hemólisis y eritropoyesis ineficaz sin ningún cambio en las reservas corporales de hierro. Por último, también se encuentran concentraciones elevadas de ferritina sin sobrecarga de hierro en el recién reconocido síndrome de hieperferritinemia hereditaria y el de catarata congénita autosómico dominante.36
La saturación de transferrina es el indicador indirecto más útil para investigar hemocromatosis ligada al HLA y otras formas de sobrecarga parenquimatosa de hierro27 [ver figura 6 y tabla 1]. Los valores de saturación de transferrina por encima del 50 porciento para las mujeres y del 60 porciento para los hombres requieren de mayor investigación. Aunque útil como prueba de escrutinio, no refleja en forma cuantitativa las reservas de hierro del organismo. La labilidad de la saturación de transferrina, en especial en la hemocromatosis ligada al HLA temprana, posee otros problemas de interpretación. La saturación de transferrina también se altera por infección, inflamación, enfermedad hepática, neoplasias, quimioterapia citotóxica y otros trastornos. Por lo tanto, existen condiciones, como la enfermedad hepática, en las que la saturación de ferritina sérica y la saturación de transferrina pueden estar elevadas sin un aumento en la reserva de hierro del organismo. La saturación de transferrina disminuye tanto en la deficiencia de hierro [ver figura 6 y tabla 1] como en la anemia por inflamación crónica,37 pero los valores de saturación de transferrina se superponen mucho en estas dos condiciones, lo que hace la medición de poca utilidad para distinguir entre ambos padecimientos.
La concentración del receptor de transferrina en plasma es una prueba nueva útil para detectar deficiencia de hierro en presencia de inflamación o infección.38 El receptor de transferrina que se encuentra en el plasma circulante es una forma trucada del receptor de transferrina celular dimérico. El receptor de transferrina soluble corresponde al dominio citoplásmico N-terminal, que al parecer se desprende por acción proteolítica de la membrana celular. Se considera que la concentración del receptor de transferrina en plasma depende de la masa total del receptor tisular. En personas normales la mayoría de los receptores celulares de transferrina se encuentran en los precursores eritroides en la médula ósea, y la mayoría de los receptores solubles provienen de estas células eritroides. En consecuencia, la concentración del receptor de transferrina en plasma está determinada principalmente por la actividad de la médula eritroide. La concentración plasmática del receptor de transferrina está disminuida cuando existe hipoplasia (o aplasia) eritroide, como en el caso de la insuficiencia renal crónica o la anemia aplástica. Por el contrario, la concentración del receptor de transferrina en plasma aumenta en pacientes con hiperplasia eritroide, esto es, en pacientes con talasemia mayor, anemia de células falciformes y otros estados hemolíticos crónicos. En ausencia de hiperplasia eritroide, un aumento en la concentración del receptor de transferrina proporciona un método útil para detectar la deficiencia de hierro tisular. La concentración del receptor de transferrina puede ser especialmente útil para distinguir la anemia por deficiencia de hierro de la anemia asociada con la inflamación crónica o la infección. Mientras que la concentración sérica de ferritina puede aumentar con niveles normales, disminuidos o ausentes de hierro en los pacientes con infección, inflamación o enfermedad hepática, la concentración del receptor de transferrina en plasma parece no afectarse por estos trastornos concomitantes. En estas circunstancias, la concentración del receptor de transferrina en plasma proporciona un indicador nuevo y aparentemente más confiable de la deficiencia de hierro.39 La concentración del receptor de transferrina en plasma no es útil para diagnosticar sobrecarga de hierro.
La excreción urinaria de hierro con agentes quelantes, como la desferroxamina o el ácido dietilenetriamino penta-acético (DPTA), constituye otro indicador del estado del hierro en el organismo,7 que es especialmente útil para evaluar la sobrecarga de hierro. Después de administrar una dosis estándar de un agente quelante, existe considerable superposición de las tasas de excreción de hierro en la orina en las personas con reservas normales y en las que tienen reservas disminuidas, de modo que esta prueba no es útil para diagnosticar deficiencia de hierro. La medición de la excreción de hierro en orina en ocasiones puede ayudar a detectar la sobrecarga del mismo, pero este indicador puede verse influenciado por la deficiencia de ácido ascórbico, infecciones, inflamación, enfermedad hepática y otros padecimientos.
La protoporfirina zinc eritrocitaria es un indicador de la cantidad de hierro disponible en la médula ósea cuando se produjeron los eritrocitos circulantes.39 El paso final en la biosíntesis del hem consiste en la inserción de un ión ferroso a una protoporfirina IX para producir el hem. En ausencia de hierro se inserta zinc, lo que origina una protoporfirina zinc, que persiste durante toda la vida del eritrocito. Como resultado, la concentración de protoporfirina zinc refleja el aporte de hierro a la médula eritroide durante las semanas previas. Como con algunos otros indicadores del estado del hierro, el aumento en la protoporfirina zinc eritrocitaria no tiene especificidad y es influenciada por varias condiciones que limitan el aporte de hierro, como la deficiencia de ácido ascórbico, la infección, la inflamación, la enfermedad hepática y las neoplasias. Los niveles de protoporfirina zinc también aumentan en muchas anemias sideroblásticas y, en especial, en la intoxicación crónica por plomo. La protoporfirina zinc eritrocitaria no es de utilidad en el diagnóstico de la sobrecarga de hierro, pero es útil como prueba de escrutinio para la intoxicación por plomo.
El examen de la sangre periférica no ayuda a detectar sobrecarga de hierro, que no produce alteraciones diagnósticas, y puede detectar cambios en la reducción del hierro corporal solo después de que la depleción de las reservas de hierro restringe el aporte de hierro para la eritropoyesis. Estas limitaciones se aplican cuando el frotis es examinado en forma visual o con el uso de instrumentos para determinar concentración de hemoglobina, hematocrito, índices eritrocitarios, distribución del volumen de los eritrocitos y cuenta de reticulocitos.8 Los cambios observados en la sangre periférica no son específicos de deficiencia de hierro y pueden ser también causados por cualquier trastorno que afecte la síntesis de hemoglobina, como la talasemia, la infección, la inflamación y las neoplasias.
Exceso de hierro
El exceso de hierro se origina de un aumento constante en el aporte de hierro en relación con los requerimientos, y causa cambios característicos en la función, transporte y almacén del hierro [ver figura 6 y tabla 1]. La cantidad de hierro corporal suele controlarse en condiciones normales por regulación en la absorción dietética de hierro. Se desarrolla exceso de hierro en condiciones que modifican o superan la regulación de la absorción del ión en el intestino [ver tabla 2]. Debido a que los humanos no tienen una forma fisiológica de eliminar el exceso de hierro, cualquier incremento persistente en la ingesta puede eventualmente causar exceso de hierro. Independientemente de la causa del acúmulo del hierro, cuando la cantidad del mismo excede la capacidad del organismo para secuestrar el exceso se desarrolla daño tisular potencialmente letal. Las características precisas de las consecuencias patológicas de la sobrecarga de hierro son resultado, en parte, de la magnitud de la carga de hierro corporal, la velocidad a la que ha ocurrido el aumento, la distribución específica del exceso del ión entre los sitios de almacén en macrófagos y los depósitos, potencialmente más dañinos, en células parenquimatosas, y la coexistencia de condiciones que pueden disminuir (v.gr., deficiencia de ácido ascórbico) o pueden empeorar (v.gr., uso de alcohol o hepatitis) la evolución. Las consecuencias más comunes de la sobrecarga de hierro incluyen enfermedad hepática, enfermedad pancreática asociada con diabetes mellitus, trastornos endócrinos asociados con insuficiencia gonadal, disfunción cardiaca, artropatía y, en ocasiones, alteraciones neurológicas y psicológicas.
|
|
* Puede tener un componente genético |
EXCESO DE HIERRO CAUSADO POR MAYOR ABSORCION
Puede ocurrir mayor absorción de hierro por un control anormal en su absorción aún cuando la dieta contenga cantidades normales de hierro biodisponible, como en la hemocromatosis hereditaria (asociada al HLA), o por una dieta con tanto exceso de hierro biodisponible que supera los mecanismos de control, como en el exceso dietético de hierro que ocurre en africanos.
Hemocromatosis hereditaria (asociada al HLA)
La hemocromatosis hereditaria es la causa más común de sobrecarga de hierro en los Estados Unidos. Hasta el 10 porciento de la población es heterocigota para esta condición, y se cree que el estado homocigoto afecta hasta al 0.5 porciento de la población, o alrededor de un millón de personas.27,40 En este trastorno el defecto genético depende de una alteración en el control de la absorción de hierro que causa un aumento inapropiado en la captación del mismo, con incremento progresivo en las reservas corporales. El exceso de hierro tiene un patrón de depósito principalmente parenquimatoso al inicio, sobre todo en los hepatocitos, y subsecuentemente el hierro se acumula en páncreas, corazón y otros órganos.27 Es característico que el hierro de los macrófagos en la médula ósea puede ser normal o incluso estar disminuido a pesar de un depósito parenquimatoso importante [ver figura 6 y tabla 1]. En los homocigotos que presentan hemocromatosis ligada al HLA en la edad media o después, la tétrada clásica de signos clínicos consiste en enfermedad hepática, diabetes mellitus, pigmentación cutánea y falla gonadal.41 Se desarrolla insuficiencia cardiaca en alrededor del 10 a 15 porciento de los homocigotos no tratados. Con menos frecuencia, los pacientes más jóvenes con hemocromatosis presentan alteraciones cardiacas y falla gonadal como signos clínicos iniciales.42 Las reservas de hierro corporal suelen haber aumentado de la cantidad normal de 1 g o menos a 15 o 20 g o más cuando ocurren los síntomas de daño parenquimatoso, por lo general en la edad media o tardía.43 Los aumentos adicionales en el hierro corporal pueden ser fatales, aunque algunos pacientes son capaces de tolerar una reserva total de hierro de hasta 40 a 50 g. Los factores ambientales, incluyendo el contenido de hierro de la dieta y el uso de alcohol, pueden influir mucho en la velocidad y severidad del daño orgánico.44 En los heterocigotos la hemocromatosis ligada al HLA se expresa en forma incompleta, y solo alrededor del 25 porciento de estas personas sufren un aumento leve y aparentemente inofensivo en las reservas de hierro, con cantidades totales de 4 o 5 g.29 Se ha reportado un exceso más importante de hierro en los pacientes heterocigotos para hemocromatosis con esferocitosis hereditaria coexistente, pero el efecto de un alelo para hemocromatosis sobre la reserva de hierro es incierta en la anemia sideroblástica idiopática y en otros trastornos hereditarios o adquiridos.
La detección de la hemocromatosis ligada al HLA aún depende de la expresión fenotípica de la enfermedad. Aún no se ha identificado el gen responsable, y el análisis directo del ADN genómico no es factible como medio para identificar a los homocigotos. El locus de la hemocromatosis está unido en forma estrecha al locus del HLA-A en el brazo corto del cromosoma 6. Por lo tanto, si se identifica un individuo afectado (probando), puede usarse la tipificación del HLA para clasificar los genotipos de los familiares en primer grado como normales, heterocigotos u homocigotos. En cambio, la tipificación del HLA no es útil como instrumento de escrutinio de población porque los haplotipos HLA específicos ligados al locus de la hemocromatosis pueden ser diferentes en cada familia. Para propósitos de escrutinio, la medición del hierro en plasma, la saturación de transferrina y la ferritina en plasma27 proporcionan el mejor medio indirecto para estudiar a personas que son homocigotas para la hemocromatosis ligada al HLA, aunque debe reconocerse que la ferritina en plasma puede ser normal en un pequeño número de pacientes con hemocromatosis ligada al HLA.45 Si cualquiera de estas medidas es anormal, está indicada una evaluación posterior. La biopsia hepática permite un diagnóstico definido de la hemocromatosis ligada al HLA.27 La evaluación debe incluir determinación cuantitativa de la concentración de hierro no hem, evaluación histoquímica del patrón de depósito de hierro y evaluación patológica de la lesión tisular. El cálculo del índice hepático de hierro (la concentración hepática de hierro [expresada como µmol Fe/g de tejido hepático en peso seco] dividida entre la edad del paciente [expresada en años])46 puede ser útil para distinguir a los homocigotos para la hemocromatosis asociada al HLA de los heterocigotos o de los pacientes con aumento en el hierro corporal asociado con enfermedad hepática crónica (por lo general alcohólica). Es típico encontrar aumento en el índice de hierro hepático a 2.0 o más en las personas con hemocromatosis asociada al HLA homocigotas, mientras que el índice suele ser menor de 1.8 en los heterocigotos y menos de 1.4 en los pacientes con hepatopatía alcohólica crónica. En ciertos casos puede usarse la flebotomía cuantitativa para establecer el diagnóstico de estado homocigoto en forma retrospectiva si se eliminan más de 5 g de hierro. Siempre que se establezca el diagnóstico de hemocromatosis es obligatorio evaluar a los familiares con riesgo de la enfermedad. El escrutinio debe incluir no solo a los hermanos, sino también a los padres e hijos por la posibilidad de parejas homocigotas-heterocigotas. En estos estudios familiares puede ser útil la tipificación del HLA.
Tratamiento de la hemocromatosis asociada al HLA El tratamiento de elección para la hemocromatosis asociada al HLA es la flebotomía para reducir y mantener el hierro corporal en niveles normales o casi normales. En los pacientes con hemocromatosis asociada al HLA e insuficiencia cardiaca se ha sugerido el uso de flebotomía y tratamiento quelante. El tratamiento con flebotomías debe iniciarse en cuanto se hace el diagnóstico de estado homocigoto para la enfermedad, su postergación solo aumenta el riesgo de daño orgánico por exceso de hierro. El programa de flebotomía debe eliminar 500 ml de sangre (conteniendo 200 a 250 mg de hierro) una vez por semana o, en pacientes con exceso importante, dos veces por semana,47 hasta que el paciente tenga deficiencia de hierro. Antes de cada flebotomía debe medirse el hematocrito o la hemoglobina. Al principio estos deben disminuir alrededor de 10 porciento de sus valores iniciales, pero después pueden aumentar al aumentar la velocidad de la eritropoyesis para satisfacer las demandas de la flebotomía. Debe medirse en forma regular la ferritina, el hierro y la saturación de transferrina, para vigilar el progreso en la eliminación del hierro. Al eliminar hierro la concentración de ferritina en plasma disminuirá en forma progresiva, pero el hierro y la saturación de transferrina permanecerán elevados hasta que las reservas estén casi agotadas. Por último, cuando se ha eliminado todo el hierro acumulado la ferritina disminuirá a menos de 12 µg/L, el hierro y la saturación de transferrina caerán y la concentración de hemoglobina disminuirá a menos de 10 g/dl durante dos semanas sin otra flebotomía. Para ilustrar qué tanto tiempo puede requerirse el tratamiento, si la reserva inicial de hierro es de 25 g, su eliminación total con flebotomías semanales puede necesitar de dos años o más. Después de haber terminado la eliminación del hierro se requiere un programa de por vida de flebotomías de mantenimiento para evitar su reacúmulo. Típicamente se necesitan flebotomías de 500 ml cada tres a cuatro meses. El objetivo de la fletobomía de mantenimiento debe ser mantener una saturación de transferrina normal con una concentración de ferritina en plasma de menos de alrededor de 50 µg/L.
La concentración de hierro no hem hepático es un factor de riesgo importante para el desarrollo de cirrosis en la hemocromatosis asociada al HLA. En ausencia de hepatopatía alcohólica, no suele desarrollarse fibrosis o cirrosis hasta que el hierro almacenado en el hígado alcanza una concentración de 400 µmol Fe/g de hígado en peso seco (rango normal: alrededor de 3 a 30 µmol Fe/g de hígado en peso seco).46 Si el tratamiento con flebotomía elimina la carga de hierro antes que se desarrolle cirrosis la esperanza de vida para el paciente es normal.43 Si por el contrario ya existe cirrosis, el riesgo de carcinoma hepatocelular aumenta en más de 200 veces. En la hemocromatosis asociada al HLA se desarrollan hepatomas casi en forma exclusiva en los pacientes con cirrosis hepática, siendo la causa de muerte en el 20 a 30 porciento de estos pacientes, incluso después de la eliminación exitosa del exceso de hierro. En la actualidad la cirrosis y el carcinoma hepatocelular son las dos principales causas de muerte en la hemocromatosis asociada al HLA.48 Casi siempre está indicado el tratamiento con flebotomías para los pacientes con hemocromatosis asociada al HLA, incluso si ya existe cirrosis o daño orgánico, porque puede detenerse la progresión de la enfermedad e incluso mejorar parte de la disfunción orgánica.
Anemias con exceso de hierro
En los pacientes con el grupo heterogéneo de padecimientos clasificados como anemias con exceso de hierro, puede desarrollarse sobrecarga severa de hierro como resultado de una mayor absorción gastrointestinal del mismo.49 Cualquier transfusión de eritrocitos que reciban estos pacientes contribuye al acúmulo del hierro. Estas anemias por acúmulo de hierro pueden ser heredadas o adquiridas, pero todas parecen tener en común una importante hiperplasia eritroide con eritropoyesis ineficaz marcada. Las anemias con sobrecarga de hierro incluyen anemia diseritropoyética congénita, deficiencia de piruvato cinasa, talasemia mayor (anemia de Cooley) y talasemia intermedia, talasemia-§ con hemoglobina E, diversas formas de anemia sideroblástica, algunas anemias mielodispásicas y varios otros trastornos anémicos en los que se altera la incorporación del hierro a la hemoglobina. Debido a que la magnitud de la eritropoyesis ineficaz, y no la severidad de la anemia, parece determinar la velocidad de acúmulo de hierro, puede desarrollarse sobrecarga severa de hierro en pacientes con anemia solo leve. Las manifestaciones clínicas y la patología que puede desarrollarse en pacientes con anemias con exceso de hierro es semejante a las de la hemocromatosis asociada al HLA, incluyendo enfermedad hepática, diabetes mellitus, trastornos endócrinos y disfunción cardiaca.
Otras formas de exceso de hierro causadas por aumento en la absorción
Algunos pacientes con hepatopatía crónica, incluyendo los que tienen cirrosis alcohólica y con derivación portocava,46 pueden sufrir grados leves o moderados de exceso de hierro como resultado de mayor absorción del hierro de la dieta. Los mecanismos responsables de la mayor captación gastrointestinal de hierro no se han identificado, aunque se han propuesto como factores etiológicos a la eritropoyesis ineficaz y a la hiperferremia asociada con alteraciones sideroblásticas y del folato inducidas por alcohol.7 Las reservas corporales de hierro están aumentadas solo un poco, típicamente a 2 a 4 g. El depósito de hierro predomina en las células de Kupffer, más que en los hepatocitos, como en la hemocromatosis asociada al HLA. Como ya se mencionó, la distinción entre la sobrecarga de hierro en los pacientes con hepatopatía crónica y la de los homocigotos para la hemocromatosis asociada al HLA casi siempre puede hacerse por determinación cuantitativa de hierro en una muestra de hígado obtenida por biopsia calculando el índice de hierro hepático.
Los pacientes sintomáticos con porfiria cutánea tarda, una porfiria hepática, suelen tener un aumento modesto en el hierro corporal que casi siempre se debe a mayor absorción gastrointestinal. En la porfiria cutánea tarda el hígado produce porfirinas fotosensibilizantes que después circulan hacia la piel.8 La causa del exceso de hierro se desconoce, pero la eliminación del hierro por flebotomía o tratamiento quelante casi siempre causa mejoría clínica y bioquímica importante. Recientemente se ha reconocido a la aceruloplasminemia como un trastorno autosómico recesivo del metabolismo del hierro que causa acúmulo del mismo tanto en el hígado como en el cerebro, manifestándose como un padecimiento neurológico en las etapas tardías de la vida.50,51 Se han descubierto otros defectos congénitos raros asociados con sobrecarga de hierro. En los pacientes con atransferrinemia prácticamente no existe transferrina. La absorción gastrointestinal de hierro está aumentada, pero debido a que no existen mecanismos fisiológicos para su transporte, casi nada del hierro absorbido puede ser captado por las células eritroides en desarrollo. El hierro absorbido se acumula entonces en el hígado, páncreas, corazón, tiroides y riñones,52 con casi nada de hierro depositado en la médula ósea o el bazo. Se han encontrado depósitos marcado de hierro hepático y fibrosis asociados con un defecto congénito que parece consistir en una captación defectuosa del hierro por los precursores eritroides, pero la fisiopatología de este trastorno aún no se conoce.53
Se ha descrito exceso dietético de hierro en africanos en por lo menos nueve países en la región sub-Sahara de Africa en asociación con una ingesta muy abundante de hierro por una bebida fermentada de maíz preparada en casa en tambos de acero.7 El acúmulo de hierro puede ser tan importante como el encontrado en la hemocromatosis asociada al HLA, y puede desarrollarse enfermedad hepática (con cirrosis y hepatoma), daño pancreático (con diabetes mellitus), trastornos endócrinos y disfunción cardiaca. Aunque durante mucho tiempo se consideró que la mayor ingesta en la dieta era la única causa de la mayor absorción de hierro en este padecimiento, los análisis de familias han sugerido que puede participar un gen no asociado al HLA,54 que puede ser común en poblaciones con ancestros africanos. Se ha demostrado sobrecarga severa de hierro en pacientes de los Estados Unidos con ancestros africanos,55 pero la relación con el exceso de hierro dietético de los africanos aún es motivo de investigación.
Sin duda la ingestión medicinal de hierro puede contribuir al acúmulo del mismo en los pacientes con estos padecimientos, pero aún no se tiene certeza del grado en el que el hierro administrado por vía oral puede aumentar las reservas corporales de hierro en los individuos normales. Aunque algunos reportes de casos han descrito exceso de hierro en pacientes que toman hierro medicinal por periodos prolongados, no puede excluirse en estos pacientes la participación potencial de un alelo no reconocido para la hemocromatosis asociada al HLA, ya que el gen aún no se identifica.
EXCESO DE HIERRO POR TRANSFUSIONES Y ADMINISTRACION PARENTERAL
Un programa adecuado de transfusiones puede mantener la vida en pacientes con anemia severa crónica y refractaria, pero el tratamiento transfusional aislado produce acúmulo progresivo por el hierro contenido en los eritrocitos transfundidos.56 El acúmulo de hierro por transfusiones ocurre al inicio sobre todo en los macrófagos, seguido de redistribución a los tejidos parenquimatosos [ver figura 6]. En los pacientes con anemias congénitas severas, como la talasemia mayor y el síndrome de Blackfan-Diamond, las transfusiones regulares pueden prevenir la muerte por anemia en la infancia y permitir un crecimiento y desarrollo normal durante la niñez. Las anemias adquiridas dependientes de transfusión, como la anemia aplástica, la aplasia pura de eritrocitos, los trastornos hipoplásicos o mielodisplásicos y otros padecimientos, pueden causar exceso de hierro corporal. Si las anemias dependientes de transfusión incluyen hiperplasia eritroide con eritropoyesis ineficaz, puede agregarse mayor absorción gastrointestinal de hierro para empeorar el acúmulo. En estos casos debe minimizarse la captación de hierro de la dieta por medio de un programa adecuado de transfusiones. Aunque no dependen de transfusiones, los pacientes con trastornos falciformes, como la anemia de células falciformes y la talasemia-§ con células falciformes, pueden adquirir una cantidad considerable de hierro por transfusiones crónicas para prevenir eventos cerebrovasculares, crisis dolorosas y otras complicaciones recurrentes. Debido a que el humano no tiene sistemas fisiológicos para eliminar el exceso de hierro, el hierro contenido en los eritrocitos transfundidos se acumula en forma progresiva y llega a dañar al hígado, corazón, páncreas y otros órganos. Suele ocurrir la muerte por insuficiencia cardiaca.57 En los pacientes más jóvenes la carga de hierro causa falla en el crecimiento y, en la adolescencia, retraso o ausencia de la maduración sexual. El hierro medicinal parenteral puede aumentar el exceso de hierro en los pacientes con anemias microcíticas refractarias que son diagnosticados en forma equívoca como con deficiencia de hierro.
Tratamiento del exceso de hierro por transfusiones
Con cada unidad transfundida de eritrocitos se agregan a la reserva corporal alrededor de 200 a 250 mg de hierro. La mayoría de los pacientes dependientes de transfusión requieren de 200 a 300 ml/kg/año de sangre. Por ejemplo, un adulto de 70 kg requiere alrededor de dos a tres unidades de sangre cada tres o cuatro semanas, lo que agrega de 6 a 10 g de hierro por día. La severidad de la toxicidad por hierro parece relacionarse con la magnitud de la carga de hierro en el organismo. Casi todos los pacientes que han sido tratados solo con transfusión y han recibido 100 o más unidades de sangre58 (alrededor de 20 a 25 g de hierro) han desarrollado depósitos de hierro en el corazón, con frecuencia asociados con signos de daño hepático, pancreático y endócrino.
Para los pacientes que son dependientes de transfusión o tienen anemia severa, la única manera de superar la limitación fisiológica que evita la eliminación de hierro es con el tratamiento con un agente quelante capaz de formar complejos con el hierro y permitir su excreción. El único agente quelante disponible en la actualidad para uso clínico es un sideróforo bacteriano que comenzó a usarse hace tres décadas, la desferroxamina B, un ácido trihidroxámico producido por Streptomyces pilosus. En la actualidad existen estudios clínicos que han demostrado la eficacia de la quelación de hierro como un enfoque terapéutico a su exceso,59 demostrando que la quelación regular de este elemento puede disminuir su exceso corporal, reducir la disfunción orgánica y mejorar la supervivencia.30,60 Aunque en la actualidad se han reconocido diversos efectos adversos tóxicos, en especial con el tratamiento intensivo, la desferroxamina es un medicamento muy seguro, incluso cuando se ha usado practicamente de por vida en algunos pacientes.
El tratamiento de quelación del hierro debe comenzarse pronto para evitar el acúmulo de cantidades tóxicas de hierro en los tejidos susceptibles y para mantener las reservas corporales de hierro en concentraciones asociadas con un riesgo bajo de muerte temprana y complicaciones clínicas.30,60 Mientras más se retrase el tratamiento quelante mayor será el riesgo de toxicidad por hierro. Debido a que la desferroxamina se absorbe poco después de la administración oral, el medicamento debe administrarse por vía parenteral para ser activo. Además, la desferroxamina se elimina con tanta rapidez de la circulación que se requieren infusiones prolongadas para tener un efecto terapéutico.61 Como resultado, para que sea útil en los pacientes con exceso de hierro, debe administrarse por infusión subcutánea o intravenosa lenta durante nueve a 12 horas de cada día por lo menos durante cinco días de la semana. En los pacientes con talasemia mayor y otras anemias refractarias congénitas que son dependientes de transfusión desde la infancia temprana, es mejor comenzar el tratamiento quelante después de 10 a 20 transfusiones, por lo general alrededor de los tres años de edad, y administrar infusión subcutánea lenta en dosis no mayores de 25 mg/kg de peso/día para minimizar el riesgo de retraso en el crecimiento.62 En los pacientes ancianos y adultos con anemias refractarias adquiridas que requieren de transfusión regular y en los que tienen enfermedad de células falciformes que son transfundidos en forma crónica para prevenir complicaciones también parece prudente el tratamiento temprano, comenzando después de la transfusión de 10 a 20 unidades de sangre. La dosis habitual de desferroxamina en estos pacientes no debe ser mayor de 50 mg/kg de peso corporal/día, administrados en nueve a 12 horas por infusión subcutánea lenta por lo menos cinco días de la semana. En algunos pacientes que no son capaces de tolerar el dolor y molestia local de la infusión subcutánea o que requieren de la reducción rápida del exceso de hierro puede administrarse desferroxamina intravenosa a través de un reservorio implantable con acceso venoso.63 Puede ser difícil cumplir con estos esquemas casi diarios de infusión prolongada subcutánea o intravenosa, y la falta de constancia es el principal obstáculo para que el tratamiento sea eficaz.64 La administración de ácido ascórbico puede aumentar la excreción de hierro inducida por desferroxamina, pero se asocia con el riesgo de redistribución interna de hierro desde sitios de almacén relativamente benignos en los macrófagos hasta reservorios potencialmente tóxicos en las células parenquimatosas. Aunque la evidencia es anecdótica,65 las dosis grandes de ácido ascórbico deben considerarse como peligrosas en los pacientes con sobrecarga de hierro. Por lo general la desferroxamina es segura y no tóxica en el paciente con exceso de hierro, pero se han reportado complicaciones sistémicas, incluyendo reacciones anafilactoides alérgicas, complicaciones infecciosas, trastornos visuales y disfunción auditiva,66 y retraso del crecimiento.62 Como resultado, debe incluirse la evaluación regular de toxicidad medicamentosa en el tratamiento de cualquier paciente que reciba desferroxamina, incluyendo audiometrías anuales, examen de retina y evaluación del crecimiento en niños y adolescentes.62 El riesgo de muchas de estas complicaciones puede reducirse ajustando la dosis de desferroxamina según la magnitud del exceso de hierro.67 En la actualidad se están desarrollando agentes quelantes activos por vía oral, pero ninguno está clínicamente disponible aún.68,69
OTRAS FORMAS DE EXCESO DE HIERRO
Se ha detectado exceso de hierro perinatal en asociación con algunas alteraciones metabólicas raras del neonato, incluyendo tirosinemia hereditaria (hipermetioninemia), síndrome cerebrohepatorrenal o síndrome de Zellweger, y hemocromatosis perinatal, también conocida como hemocromatosis neonatal o enfermedad por almacenamiento de hierro neonatal.70 El secuestro de hierro en otros trastornos raros produce varios patrones de depósito de hierro localizado en la hemosiderosis pulmonar idiopática, la hemosiderosis renal y la enfermedad de Hallervorden-Spatz.71
Deficiencia de hierro
La deficiencia de hierro se refiere a condiciones en las que el hierro corporal está disminuido debido a un aumento sostenido en los requerimientos de hierro en comparación con su aporte. Los requerimientos de hierro incluyen no solo el hierro para restablecer las pérdidas fisiológicas y satisfacer las necesidades durante el crecimiento y el embarazo, pero tambin el que suple las pérdidas patológicas. La pérdida diaria fisiológica de hierro en el varón es un poco menor de 1.0 mg/día, y en la mujer normal que menstrúa de 1.5 mg/día.7 Pueden identificarse estadios secuenciales en la reducción del hierro corporal [ver figura 6 y tabla 1]. La reducción en las reservas de hierro sin un cambio en la cantidad de compuestos de hierro funcionales se denomina como reducción en la reserva de hierro. Cuando las reservas se agotan, se dice que los pacientes tienen depleción de hierro. La mayor disminución en el hierro corporal causa menor producción de hemoglobina y de otros compuestos funcionales que contienen hierro, estadio denominado eritropoyesis deficiente en hierro. La reducción aún mayor en el hierro corporal produce anemia por deficiencia de hierro. En todo el mundo, la deficiencia de hierro es la causa más común de anemia.39 Casi la tercera parte de la población mundial de alrededor de cinco billones tiene anemia, y la anemia por deficiencia de hierro causa o contribuye a la anemia en por lo menos 500 a 600 millones de personas.72
MANIFESTACIONES CLINICAS DE LA DEFICIENCIA DE HIERRO
Desde el punto de vista clínico los pacientes con deficiencia de hierro pueden estar asintomáticos y detectarse solo por resultados anormales en las pruebas de laboratorio. Otros pacientes pueden solicitar atención médica por las manifestaciones del trastorno subyacente que causó la deficiencia de hierro, pero pueden no tener datos relacionados con esta última. Un tercer grupo de enfermos puede sufrir signos y síntomas comunes a todas las anemias, como debilidad, mareo, fatiga fácil, palidez, irritabilidad y otras molestias indefinidas e inespecíficas. Por último, algunos pacientes presentan uno o más de los pocos signos y síntomas que se piensa son muy específicos de la deficiencia de hierro (escleras azules, coiloniquia y pagofagia).1 La deficiencia de hierro puede asociarse también con signos y síntomas que no se relacionan con anemia,73 como estomatitis angular, glositis, estenosis o pliegues esofágicos poscricoideos y atrofia gástrica. El síndrome de Plummer-Vinson, también conocido como síndrome de Paterson-Kelly,7 se manifiesta por glositis, ardor o sensación quemante en la boca, disfagia y deficiencia de hierro. La pagofagia, o pica con hielo, parece ser un síntoma muy específico de deficiencia de hierro y desaparece poco después de que se inicia el tratamiento.74 Otros tipos de pica pueden acompañar a la deficiencia de hierro, pero ninguno es tan específico como la pagofagia. En la coiloniquia, la porción distal de las uñas de los dedos adoptan forma de cuchara, y las uñas son delgadas y brillantes, quizá como consecuencia de trastornos en el crecimiento del lecho epitelial ungueal. Aunque se piensa que la coiloniquia es muy específica de la deficiencia de hierro, este signo ocurre en un número muy pequeño de pacientes. Las escleras con coloración azul se han descrito como un indicador muy sensible y específico de deficiencia de hierro.75 Esta coloración azulosa parece deberse a adelgazamiento de la esclerótica, quizá como consecuencia de una menor síntesis de colágena por la deficiencia de hierro. La deficiencia de hierro puede producir otras manifestaciones clínicas que no se relacionan con la anemia y que parecen ser causadas por la depleción de compuestos funcionales de hierro en los tejidos no eritroides. Las consecuencias no hematológicas de la falta de hierro incluyen menor tolerancia al ejercicio y desempeño laboral y deterioro en la inmunidad y resistencia a la infección. En los niños la deficiencia de hierro afecta en forma adversa el crecimiento, el desarrollo motor y las funciones cognoscitivas y de comportamiento. Estas alteraciones pueden no ser reversibles con el tratamiento posterior.39,73,76-78
CAUSAS DE DEFICIENCIA DE HIERRO
La pérdida de sangre es la causa más común de aumento en el requerimiento de hierro que origina deficiencia del mismo [ver tabla 3]. En los hombres y las mujeres posmenopáusicas, la deficiencia de hierro casi siempre es resultado de pérdida gastrointestinal.39 En las mujeres menstruando la pérdida genitourinaria suele ser la causa del aumento en los requerimientos de hierro. Los anticonceptivos orales pueden disminuir la pérdida menstrual de sangre, pero los dispositivos intrauterinos la aumentan. Deben considerarse también otras causas de hemorragia genitourinaria o respiratoria [ver tabla 3]. Para los donadores de sangre, cada donación significa una prdida de 200 a 250 mg de hierro. Durante los periodos de crecimiento en la infancia, niñez y adolescencia, los requerimientos de hierro pueden superar el aporte a partir de las dietas y reservas.73 El embarazo aumenta las necesidades de hierro y, sin hierro suplementario, causa una pérdida neta equivalente a alrededor de 1,200 a 1,500 ml de sangre. Después del parto, si el niño es alimentado al seno materno, la lactación aumenta los requerimientos de hierro en alrededor de 0.5 a 1.0 mg por día.
|
||
|
El aporte insuficiente de hierro puede contribuir al desarrollo de deficiencia de hierro. En los niños y mujeres con requerimientos altos de hierro, las dietas que contienen cantidades inadecuadas de hierro biodisponible pueden aumentar el riesgo de deficiencia. En los niños mayores, varones y mujeres posmenopáusicas, el aporte escaso de hierro en la dieta casi nunca es el único factor responsable de la deficiencia, por lo que deben buscarse otros factores etiológicos, en especial pérdida de sangre.
La menor absorción de hierro es una causa poco común de deficiencia. En algunos pacientes la malabsorción intestinal de hierro es solo un aspecto más de la malabsorción generalizada [ver tabla 3]. La cirugía gástrica, en especial la resección gástrica parcial o total o la gastroenterostomía para derivación del duodeno, puede causar deficiencia de hierro. Aunque la absorción de hierro en la dieta puede ser pobre, las sales terapéuticas se absorben bien, y la deficiencia puede corregirse con facilidad.
El riesgo de deficiencia de hierro es especialmente alto cuando los requerimientos aumentan y el aporte es inadecuado. Por ejemplo, los infantes alimentados con leche de vaca suelen tener deficiencia de hierro por la combinación de mayor pérdida por hemorragia gastrointestinal inducida por la leche de vaca y por las pequeñas cantidades de hierro biodisponible que existen en ésta.73 Las mujeres con requerimientos elevados de hierro por la menstruación suelen consumir dietas que tienen poco hierro biodisponible y con inhibidores de la absorción de hierro, como el calcio.
DIAGNOSTICO DE LA DEFICIENCIA DE HIERRO
La anemia por deficiencia de hierro es la única anemia microcítica hipocrómica asociada con ausencia en las reservas de hierro. En las otras anemias microcíticas hipocrómicas las reservas de hierro en la médula son normales o están aumentadas. Pueden usarse medidas indirectas del hierro corporal para identificar una secuencia característica de cambios que ocurren al disminuir el hierro corporal desde los niveles de reserva normal hasta los encontrados en la anemia por deficiencia de hierro [ver figura 6 y tabla 1]. La concentración de ferritina en plasma es la prueba más útil para la detección de la deficiencia de hierro porque las concentraciones de ferritina en plasma disminuyen al reducirse las reservas corporales.4 Una concentración de ferritina en plasma por debajo de 12 µg/L es virtualmente diagnóstica de ausencia de reservas de hierro. El hipotiroidismo y la deficiencia de ácido ascórbico pueden disminuir la concentración de ferritina en plasma en forma independiente de la reducción en el hierro corporal, pero estos trastornos no suelen constituir un problema diagnóstico.79 Por el contrario, la concentración normal de ferritina en plasma no confirma la presencia de reservas de hierro porque esta proteína puede aumentar en forma independiente por infecciones, inflamación, enfermedad hepática, neoplasias y otros trastornos. En estos casos la medición de la concentración del receptor de transferrina en plasma proporciona una manera de distinguir entre la anemia por deficiencia de hierro y la anemia asociada con trastornos inflamatorios crónicos.38 A diferencia de la concentración plasmática de ferritina, la concentración de receptor de transferrina en plasma no parece afectarse por infecciones, inflamación, enfermedad hepática o neoplasias, y puede ser un medio más eficaz para detectar anemia por deficiencia de hierro. La saturación de transferrina en plasma está disminuida tanto en la deficiencia de hierro como en los estados infecciosos e inflamatorios, y es de poca ayuda práctica para distinguir entre estos trastornos. Aunque rara vez se realiza examen de la médula ósea solo para evaluar el estado del hierro, casi siempre puede verificarse el estado de deficiencia de hierro por la evaluación directa de las reservas de este elemento en la médula. Si no existen reservas de hierro se establece diagnóstico de deficiencia de hierro, si se encuentra hemosiderina, se excluye esta posibilidad. Además, en la deficiencia de hierro no existen sideroblastos en la médula, o estos están muy disminuidos, constituyendo menos del 10 porciento de los normoblastos. Una prueba terapéutica con hierro puede ser un medio eficaz de establecer el diagnóstico de deficiencia de hierro.
TRATAMIENTO DE LA DEFICIENCIA DE HIERRO
El tratamiento para la anemia por deficiencia de hierro consiste en corregir tanto la deficiencia de hemoglobina como reemplazar las reservas de hierro.39,73 Para casi todos los pacientes el hierro oral es el tratamiento de elección. El tratamiento con hierro oral es eficaz, seguro y económico. Debido al riesgo de reacciones adversas locales y sistémicas, el hierro parenteral solo debe usarse en el pequeño número de pacientes que no pueden absorber o tolerar el hierro oral o que tienen requerimientos de hierro que no pueden satisfacerse por el tratamiento oral debido a hemorragia crónica no controlable. En la anemia severa por deficiencia de hierro puede requerirse transfusión de eritrocitos para los pacientes en los que la velocidad de pérdida crónica de hierro excede la velocidad a la que puede reemplazarse el hierro con tratamiento parenteral. Aunque la mayoría de los pacientes toman el hierro oral sin dificultad, el 10 a 20 porciento sufre efectos colaterales, principalmente de tipo gastrointestinal.
La deficiencia de hierro casi siempre puede tratarse en forma eficaz, y el tratamiento oral y parenteral da resultados semejantes [ver tabla 4]. Ocurre mejoría de los síntomas en los primeros días de tratamiento. En la anemia no complicada, la respuesta hematológica inicial, que consiste en reticulocitosis leve, comienza en tres a cinco días después del inicio del tratamiento, alcanza un máximo a los ocho a 10 días y disminuye después. Después de la primera semana la concentración de hemoglobina comienza a aumentar y suele ser normal alrededor de seis semanas después. La microcitosis puede no resolverse por completo hasta después de cuatro meses. Si la deficiencia de hierro se trata con hierro oral en dosis de 200 mg/día o menos, la concentración de ferritina en plasma permanece en menos de 12 µg/L hasta que la anemia se corrija y después aumenta en forma gradual al repletarse las reservas de hierro. Si la respuesta al tratamiento con hierro no es completa y característica, estará indicado reevaluar al paciente. Uno de los problemas más comunes es confundir la anemia de una enfermedad crónica con anemia por deficiencia de hierro. La recuperación puede ser más lenta cuando coexisten padecimientos, incluyendo otras deficiencias nutricionales, enfermedad hepática o renal, infecciones, inflamación o neoplasias, o pérdida de sangre oculta continua. Si el paciente fue tratado con hierro oral, deberán revisarse la forma y dosis de hierro, la constancia al tratamiento y la posibilidad de malabsorción.
|
|
|
Bibliografía
- Harris JW, Kellermeyer RW: The Red Cell. Production, Metabolism, Destruction: Normal and Abnormal. Harvard University Press, Cambridge, Massachusetts, 1970
-
Jandl JH: Blood: Textbook of Hematology. Little, Brown &
Co, Boston, 1996
-
Bunn HF, Forget BF: Hemoglobin: Molecular, Genetic and Clinical
Aspects. WB Saunders Co, Philadelphia,1986
-
Dickerson RE, Geis I: Hemoglobin. Benjamin Cummings Publishing,
Menlo Park, California, 1983
-
Holt JM, Ackers GK: The pathway of allosteric control as
revealed by hemoglobin intermediate states. FASEB J 9:210, 1995 [PMID
7781923]
-
Huang Y, Ackers GK: Enthalpic and entropic components of
cooperativity for the partially ligated intermediates of hemoglobin support
a "symmetry rule" mechanism. Biochemistry 34:6316, 1995 [PMID
7756259]
-
Bothwell TH, Charlton RW, Cook JD, et al: Iron Metabolism
in Man. Blackwell Scientific Publications, Oxford, England, 1979
-
Brittenham GM: Disorders of iron metabolism: deficiency and
overload. Hematology: Basic Principles and Practice, 2nd ed. Hoffman R,
Benz EJ, Shattil SJ, et al, Eds. Churchill Livingstone, New York, 1995,
p 492
-
Sayers MH, English G, Finch C: Capacity of the store-regulator
in maintaining iron balance. Am J Hematol 47:194, 1994 [PMID
7942783]
-
Conrad ME, Umbreit JN, Moore EG: Iron absorption and cellular
uptake of iron. Adv Exp Med Biol 356:69, 1994 [PMID
7887247]
-
Huebers HA, Finch CA: The physiology of transferrin and transferrin
receptors. Physiol Rev 67:520, 1987 [PMID
3550839]
-
Baker EN, Lindley PF: New perspectives on the structure and
function of transferrins. J Inorg Biochem 47:147, 1992 [PMID
1431877]
-
Bailey S, Evans RW, Garratt RC, et al: Molecular structure
of serum transferrin at 3.3-A resolution. Biochemistry 27:5804, 1988
-
Aisen P: Entry of iron into cells: a new role for the transferrin
receptor in modulating iron release from transferrin. Ann Neurol 32:S62,
1992
-
Zak O, Trinder D, Aisen P: Primary receptor-recognition
site of human transferrin is in the C-terminal lobe. J. Biol Chem 269:7110,1994
[PMID
8125919]
-
Munro H: The ferritin genes: their response to iron status.
Nutr Rev 51:65, 1993
-
Broxmeyer HE, Cooper S, Levi S, et al: Mutated recombinant
human heavy-chain ferritins and myelosuppression in vitro and in vivo:
a link between ferritin ferroxidase activity and biological function. Proc
Natl Acad Sci USA 88:770, 1991 [PMID
1992468]
-
Levi S, Santambrogio P, Cozzi A, et al: The role of the L-chain
in ferritin iron incorporation: studies of homo and heteropolymers. J Mol
Biol 238:649, 1994 [PMID
8182740]
-
Kuhn LC: Molecular regulation of iron proteins (review).
Baillieres Clin Haematol 7:763, 1994
-
Klausner RD, Rouault TA, Harford JB: Regulating the fate
of mRNA: the control of cellular iron metabolism. Cell 72:19, 1993 [PMID
8380757]
-
Yu Y, Radisky E, Leibold EA: The iron-responsive element
binding protein: purification, cloning, and regulation in rat liver. J
Biol Chem 267:19005, 1992 [PMID
1527027]
-
Guo B, Brown FM, Phillips JD, et al: Characterization and
expression of iron regulatory protein 2 (IRP2): presence of multiple IRP2
transcripts regulated by intracellular iron levels. J Biol Chem 270:16529,
1995 [PMID
7622457]
-
Kennedy MC, Mende-Mueller L, Blondin GA, et al: Purification
and characterization of cytosolic aconitase from beef liver and its relationship
to the iron-responsive element binding protein. Proc Natl Acad Sci USA
89:11730, 1992 [PMID
1334546]
-
Henderson BR, Kuhn LC: Differential modulation of the RNA-binding
proteins IRP-1 and IRP-2 in response to iron. IRP-2 inactivation requires
translation of another protein. J Biol Chem 270:20509, 1995
-
Guo B, Phillips JD, Yu Y, et al: Iron regulates the intracellular
degradation of iron regulatory protein 2 by the proteasome. J Biol Chem
270:21645, 1995 [PMID
7665579]
-
Samaniego F, Chin J, Iwai K, et al: Molecular characterization
of a second iron-responsive element binding protein, iron regulatory protein
2: structure, function, and post-translational regulation. J Biol Chem
269:30904, 1994 [PMID
7983023]
-
Edwards CQ, Kushner JP: Screening for hemochromatosis. N
Engl J Med 328:1616, 1993
-
Brittenham GM, Farrell DE, Harris JW, et al: Magnetic-susceptibility
measurement of human iron stores. N Engl J Med 307:1671, 1982 [PMID
7144866]
-
Nielsen P, Fischer R, Engelhardt R, et al: Liver iron stores
in patients with secondary hemochromatosis under iron chelation therapy
with deferoxamine or deferiprone. Br J Haematol 91:827, 1995 [PMID
8547125]
-
Brittenham GM, Griffith PM, Nienhuis AW, et al: Efficacy
of deferoxamine in preventing complications of iron overload in patients
with thalassemia major. N Engl J Med 331:567, 1994
-
Bonkovsky HL, Slaker DP, Bills EB, et al: Usefulness and
limitations of laboratory and hepatic imaging studies in iron-storage disease.
Gastroenterology 99:1079, 1990 [PMID
2394329]
-
Jensen PD, Jensen FT, Christensen T, et al: Evaluation of
transfusional iron overload before and during iron chelation by magnetic
resonance imaging of the liver and determination of serum ferritin in adult
non-thalassaemic patients. Br J Haematol 89:880, 1995
-
Borgna-Pignatti C, Castriota-Scanderbeg A: Methods for evaluating
iron stores and efficacy of chelation in transfusional hemosiderosis. Haematologica
76:409, 1991 [PMID
1806447]
-
Brittenham GM, Cohen AR, McLaren CE, et al: Hepatic iron
stores and plasma ferritin concentration in patients with sickle cell anemia
and thalassemia major. Am J Hematol 42:81, 1993
-
Worwood M, Cragg SJ, Jacobs A: Binding of serum ferritin
to concanavalin A of patients with homozygous thalassemia and transfusional
overload. Br J Haematol 46:409, 1980 [PMID
7448126]
-
Girelli D, Olivieri O, De Franceschi L, et al: A linkage
between hereditary hyperferritinaemia not related to iron overload and
autosomal dominant congenital cataract. Br J Haematol 90:931, 1995 [PMID
7669675]
-
Bainton DF, Finch CA: The diagnosis of iron deficiency anemia.
Am J Med 37:62, 1964
-
Cook JD, Baynes RD, Skikne BS: The physiological significance
of circulating transferrin receptors (review). Adv Exp Med Biol 352:119,
1994 [PMID
7832041]
-
Cook JD: Iron-deficiency anaemia. Baillieres Clin Haematol
7:787, 1994
-
McLaren CE, Gordeuk VR, Looker AC, et al: Prevalence
of heterozygotes for hemochromatosis in the white population of the United
States. Blood 86:2021, 1995 [PMID
7655030]
-
Sheldon JH: Haemochromatosis. Oxford University Press, London,
1935
-
Cazzola M, Ascari E, Barosi G: Juvenile idiopathic hemochromatosis.
Hum Genet 65:149, 1983
-
Niederau C, Stremmel W, Strohmeyer GW: Clinical spectrum
and management of haemochromatosis. Baillieres Clin Haematol 7:881, 1994
[PMID
7881158]
-
Fargion S, Mandelli C, Piperno A, et al: Survival and prognostic
factors in 212 Italian patients with genetic hemochromatosis. Hepatology
15:655, 1992 [PMID
1312985]
-
Feller ER, Pout A, Wands JR, et al: Familial hemochromatosis:
physiologic studies in the precirrhotic stage of the disease. N Engl J
Med 296:1422, 1977 [PMID
194151]
-
Bassett ML, Halliday JW, Powell LW: Value of hepatic iron
measurements in early hemochromatosis and determination of the critical
iron level associated with fibrosis. Hepatology 6:24, 1986 [PMID
3943787]
-
Crosby WH: Treatment of hemochromatosis by energetic phlebotomy.
One patient's response to the letting of 55 litres of blood in 11 months.
Br J Haematol 4:82, 1958
-
Fargion S, Fracanzani AL, Piperno A, et al: Prognostic factors
for hepatocellular carcinoma in genetic hemochromatosis. Hepatology 20:1426,
1994 [PMID
7982640]
-
Pootrakul P, Kitcharoen K, Yansukon P, et al: The effect
of erythroid hyperplasia on iron balance. Blood 71:1124, 1988 [PMID
3355891]
-
Morita H, Ikeda S, Yamamoto K, et al: Hereditary ceruloplasmin
deficiency with hemosiderosis: a clinicopathological study of a Japanese
family. Ann Neurol 37:646, 1995 [PMID
7755360]
-
Harris ZL, Takahashi Y, Miyajima H, et al: Aceruloplasminemia:
molecular characterization of this disorder of iron metabolism. Proc Natl
Acad Sci USA 92:2539, 1995 [PMID
7708681]
-
Goya N, Miyazaki S, Kodate S, et al: A family of congenital
atransferrinemia. Blood 40:239, 1972 [PMID
4625559]
-
Shahidi NT, Nathan DG, Diamond LK: Iron deficiency anemia
associated with an error of iron metabolism in two siblings. J Clin Invest
43:510, 1964
-
Gordeuk VR, Mukibi J, Hasstedt SJ, et al: Iron overload
in Africa: interaction between a gene and dietary iron content. N Engl
J Med 326:95, 1992
-
Barton JC, Edwards CQ, Bertoli LF, et al: Iron overload in
African Americans. Am J Med 99:616, 1995 [PMID
7503084]
-
Gabutti V, Borgna-Pignatti C: Clinical manifestations and
therapy of transfusional haemosiderosis. Baillieres Clin Haematol 7:919,
1994 [PMID
7881160]
- Model B, Berdoukas V: The clinical approach to thalassemia. Grune & Stratton, London, 1984
- Buja LM, Roberts WC: Iron in the heart, etiology and clinical significance. Am J Med 51:209, 1971 [PMID 5095527]
-
Cohen A, Martin M, Schwartz E: Depletion of excessive liver
iron stores with desferrioxamine. Br J Haematol 58:369, 1984 [PMID
6477838]
-
Olivieri NF, Nathan DG, MacMillan JH, et al: Survival in
medically treated patients with homozygous beta-thalassemia. N Engl J Med
331:574, 1994 [PMID
8047081]
-
Propper RD, Shurin SB, Nathan DG: Reassessment of the
use of desferrioxamine B in iron overload. N Engl J Med 294:1421, 1976
[PMID
1272274]
-
Olivieri NF, Koren G, Harris J, et al: Growth failure and
bony changes induced by deferoxamine. Am J Pediatr Hematol Oncol 14:48,
1992 [PMID
1550263]
-
Cohen AR, Mizanin J, Schwartz E: Rapid removal of excessive
iron with daily, high-dose intravenous chelation therapy. J Pediatr 115:151,
1989 [PMID
2738783]
-
Fosburg M, Nathan DG: Treatment of Cooley's anemia. Blood
76:435, 1990 [PMID
2198956]
-
Cohen A, Cohen IJ, Schwartz E: Scurvy and altered iron stores
in thal assemia major. N Engl J Med 304:158, 1981 [PMID
7442735]
-
Olivieri NF, Buncic JR, Chew Eea: Visual and auditory
neurotoxicity in patients receiving subcutaneous deferoxamine infusions.
N Engl J Med 314:869, 1986 [PMID
3485251]
-
Porter JB, Jaswon MS, Huehns ER, et al: Desferrioxamine ototoxicity:
evaluation of risk factors in thalassaemic patients and guidelines for
safe dosage. Br J Haematol 73:403, 1989
-
Brittenham GM: Development of iron-chelating agents for clinical
use. Blood 80:569, 1992
-
Olivieri NF, Brittenham GM, Matsui D, et al: Iron-chelation
therapy with oral deferiprone in patients with thalassemia major. N Engl
J Med 332:918, 1995 [PMID
7877649]
-
Knisely AS: Iron and pediatric liver disease. Semin Liver
Dis 14:229, 1994
-
Gerlach M, Ben-Shachar D, Riederer P, et al: Altered brain
metabolism of iron as a cause of neurodegenerative diseases? J Neurochem
63:793, 1994
-
Cook JD, Skikne BS, Baynes RD: Iron deficiency: the global
perspective. Adv Exp Med Biol 356:219, 1994
-
Oski FA: Iron deficiency in infancy and childhood. N Engl
J Med 329:190, 1993
-
Brown WD, Dyment PG: Pagophagia and iron deficiency anemia
in adolescent girls. Pediatrics 49:766, 1972
-
Kalra L, Hamlyn AN, Jones BJ: Blue sclerae: a common sign
of iron deficiency? Lancet 2:1267, 1986
- Moffatt ME, Longstaffe S, Besant J, et al: Prevention of iron deficiency and psychomotor decline in high-risk infants through use of iron-fortified infant formula: a randomized clinical trial. J Pediatr 125:527, 1994 [PMID 7523647]
-
Walter T: Effect of iron-deficiency anaemia on cognitive
skills in infancy and childhood. Baillieres Clin Haematol 7:815, 1994
-
Yip R: Iron deficiency: contemporary scientific issues and
international programmatic approaches. J Nutr 124:1479S, 1994
-
Finch CA, Bellotti V, Stray S: Plasma ferritin determination
as a diagnostic tool. West J Med 145:657, 1986 [PMID
3541387]