Inmunología
⭳ Abrir artículo (PDF)1.3 MBEste artículo fue revisado respecto a la Edición 3/2000. Ver esa versión →
Contenido del artículo
II ANTIGENOS, ANTICUERPOS Y RECEPTORES DE CELULAS T
- Antígenos
- Anticuerpos
- CLASIFICACION DE LAS INMUNOGLOBULINAS
- DIFERENCIAS ANTIGENICAS
- ESTRUCTURA Y SECUENCIA DE LAS INMUNOGLOBULINAS
- SINTESIS DE ANTICUERPOS (IN VITRO E IN VIVO)
- INMUNOGLOBULINAS COMO RECEPTORES
- ORIGEN GENETICO DE LA DIVERSIDAD DE LOS ANTICUERPOS
- HIBRIDOMAS DE LINFOCITOS
- ANTICUERPOS MONOCLONALES HUMANOS
- Receptores de células T
- Respuesta inmune humana en modelos animales
II ANTIGENOS, ANTICUERPOS Y RECEPTORES DE CELULAS T
DR. COX THERHOST, PH.D.
DR. JOHN DAVID
Antígenos
Un antígeno es cualquier sustancia capaz de generar una respuesta inmune, esto es, de reaccionar con células T y células B para inducir la formación de anticuerpos y linfocitos sensibilizados, que reaccionan con esos anticuerpos y células una vez que son formados. Los primeros antígenos estudiados fueron diversas proteínas extrañas y de microrganismos, y sigue siendo válido que las proteínas son antigénicas en forma casi universal. Las bases de la inmunogenicidad de las proteínas se desconocen, pero es probable que se relacionen con su configuración única y estable. En la actualidad se sabe que en condiciones apropiadas un amplio rango de moléculas pueden inducir una respuesta inmune y que en situaciones anormales el organismo incluso desarrollará una respuesta inmune contra sus propios antígenos.
Las reglas que gobiernan la antigenicidad proteica se están estudiando con el uso de péptidos sintéticos seleccionados por la posición que ocupan en la estructura tridimensional de las proteínas [ver figura 1]. Casi todos los anticuerpos que aparecen en forma natural dirigidos contra proteínas son específicos de sitios determinados en la superficie de las moléculas proteicas.
Cuando los polisacáridos se acoplan con proteínas pueden inducir
la formación de anticuerpos, y algunos polisacáridos purificados
son antígenos eficaces por sí solos. Un ejemplo lo constituye el
polisacárido purificado del neumococo, que puede utilizarse como vacuna
contra la cepa de neumococo en particular de la que se obtuvo el
polisacárido. Los lipopolisacáridos, como las endoxotoxinas
bacterianas, también pueden producir una respuesta de anticuerpos. Se ha
demostrado que los anticuerpos humanos contra los polisacáridos son
sobre todo de la clase IgG, aunque también se detectan cantidades
menores de IgM e IgA. Otro grupo de moléculas antigénicas son las
glucoproteínas, ejemplificadas por las proteínas que constituyen
los grupos sanguíneos y por antígenos embrionarios, tumorales,
virales y de transplante; se sabe que la especificidad de los anticuerpos
contra algunas de estas glucoproteínas está dirigida contra la
porción de carbohidratos de las moléculas.
Aunque la mayor parte de los antígenos son macromoléculas, algunas moléculas pequeñas también son antigénicas. Se ha demostrado que la hormona polipeptídica angiotensina amida, un octapéptido, y la arginina-vasopresina, un nonapéptido, pueden actuar como antígenos completos cuando se absorben en partículas de carbón. Así mismo, ciertos antibióticos conjugados con dinitrofenol (DNF), como la gramicidina-DNF y la bacitracina-DNF, actúan como antígenos al igual que el DNF(Lys), dinitrofenol conjugado con un polipéptido que contiene siete lisinas. Además, el ácido arsalínico azo- L-tirosina inducirá hipersensibilidad tardía sin formación de anticuerpos. Estas pequeñas moléculas antigénicas no contienen grupos reactivos, y por lo tanto, parecen ser antigénicas sin unirse a macromoléculas.
Se ha aprendido mucho acerca de los requerimientos de la inmunogenicidad a través del uso de polipéptidos sintéticos específicos. Por ejemplo, la agregación de aminoácidos aromáticos (como la tirosina), convertirá polipéptidos poco antigénicos en moléculas más inmunógenas aun cuando la especificidad del anticuerpo obtenido no esté dirigida contra este aminoácido. El uso de antígenos sintéticos permitió descubrir por primera vez que la respuesta inmune se encuentra bajo control genético; este descubrimiento evolucionó a partir de la observación de que algunas cepas de animales respondían a un polímero sintético, mientras que otras cepas no lo hacían.
Los antígenos no sólo son reconocidos por anticuerpos sino también a través de receptores de células T (RCT) antígeno-específicos, de los que existen dos tipos diferentes, denominados RCT-ab y RCT-gd. Los diversos tipos de células T tienen mecanismos diferentes para reconocer los antígenos. Por ejemplo, los RCT-ab reconocen antígenos que han sido procesados por células presentadoras de antígenos (CPA) para convertirse en fragmentos peptídicos unidos a moléculas del complejo principal de histocompatibilidad (CPH) de clase I o clase II en la superficie de las CPA. En contraste, los RCT-gd parecen no requerir la presentación de antígeno por el CPH. Las células T cooperadoras solo reconocen fragmentos peptídicos unidos a moléculas del CPH de clase II.1 Las células T citotóxicas reconocen antígenos procesados virales presentados tanto en moléculas de clase I como II del CPH sobre la superficie de las células humanas.2 En general, los receptores de las células T y las inmunoglobulinas reconocen diferentes determinantes antigénicos [ver adelante, Receptores de Células T]. Recientemente se ha demostrado que los lípidos puros de micobacterias pueden presentarse como antígenos al RCT-ab por las moléculas CD1 en lugar de las moléculas clase I o II del CPH.3
HAPTENOS
Existen algunas moléculas pequeñas que por sí solas no son antigénicas pero pueden reaccionar con anticuerpos; estas moléculas se denominan haptenos. Antes que ocurra la inducción de anticuerpos específicos por el hapteno, éste debe combinarse con una sustancia inmunógena, a menudo una proteína o un polipéptido sintético, denominada portador. Por ejemplo, si el hapteno fosforilcolina se combina con gamaglobulina bovina y el conjugado restante se inyecta a un animal, se producirán anticuerpos con predominio específico para el grupo fosforilcolina [ver figura 2]. Casi cualquier molécula que no sea antigénica por sí sola (diversos péptidos, esteroides, nucleótidos, lípidos y hormonas) pueden combinarse con un transportador inmunogénico y, como parte de un conjugado, inducir la formación de anticuerpos.
Esta respuesta de inducción es la base de potentes instrumentos de investigacipen el estudio de la respuesta inmune. Así mismo, es la base de muchos radioinmunoanálisis que se han convertido en auxiliares de diagnóstico útiles en medicina clínica. Además, la mayor parte de las reacciones por fármacos se producen por el hecho de que el medicamento agresor actúa como hapteno y se une en forma covalente con una proteína del propio individuo; tal vez el ejemplo más notable sea la unión covalente de la penicilina con proteínas séricas, que causa la reacción a la penicilina. La sensibilidad por contacto, incluyendo la inducida por metales, hiedra venenosa, roble venenoso, zumaque venenoso y detergentes, también es consecuencia de una molécula agresora que actúa como hapteno, que suele unirse a las proteínas de la piel.
Los anticuerpos contra haptenos de ácidos nucleicos pueden elaborarse en el laboratorio, pero esto ocurre rara vez in vitro. En pacientes con lupus eritematoso generalizado, enfermedad de Crohn o esclerosis lateral aminotrófica se producen anticuerpos antiADN y antiARN.
ADYUVANTES
Los adyuvantes son sustancias capaces de aumentar la inmunogenicidad de los antígenos. Muchos productos microbianos se han empleado como adyuvantes,4 incluyendo sustancias de Mycobacterium tuberculosis, bacilo de Calmette-Guérin (BCG), Corynebacterium parvum, Brucella abortus y Bordetella pertussis, así como toxoides de Vibrio cholerae y Clostridium tetani. Se han derivado adyuvantes del virus vaccinia y de otros poxvirus, BCG y Salmonella, cada uno transfectado con genes para un antígeno de interés. Otros son derivados de lipopolisacáridos, como el lípido monofosforil A. El adyuvante completo de Freund, que consiste en micobacterias muertas en aceite emulsificado con un antígeno en solución acuosa, se usa en la actualidad con poca frecuencia porque en general causa una intensa reacción inflamatoria local. Otros adyuvantes incluyen extractos de jabón del árbol de tejo (Quillaja saponaria), polímeros (v.gr., inulina), péptidos complejos y diversas citocinas. Por ejemplo, la interleucina-12 (IL-12) parece actuar como un promotor intenso de las reacciones inmunológicas mediadas por células en los ratones.5 La sal de aluminio alumbre también está autorizada para usarse como adyuvante en humanos.
Los liposomas también pueden actuar como adyuvantes. Las proteínas de membrana, como las proteínas de la pared viral insertadas en liposomas, son más antigénicas que aquellas solubilizadas en detergentes. En algunos casos se ha demostrado que los liposomas aumentan la respuesta inmune contra antígenos pequeños, lo cual indica que los liposomas pueden utilizarse en la elaboración de vacunas. En la práctica clínica, los adyuvantes se utilizan como complementos de algunas inmunizaciones, como la del tétanos, y son potencialmente importantes en el tratamiento antiparasitario y antitumoral.
RESPUESTA A ANTIGENOS
La respuesta del organismo a la exposición a antígenos depende de la vía de administración y de la dosis. En términos generales, los antígenos que entran al organismo por vía intravenosa viajan con rapidez al hígado, y la producción de anticuerpos es rápida. Sin embargo, la vía I.V. también facilita la inmunosupresión o la tolerancia, en especial cuando el antígeno es soluble y no agregado. Los antígenos que se administran por vía subcutánea o intradérmica suelen localizarse inicialmente en los ganglios linfáticos que drenan el sitio de la inyección, lo que favorece la inducción de inmunidad o la hipersensibilidad modulada por células. Los antígenos inhalados también favorecen la sensibilización local y el desarrollo de hipersensibilidad inmediata e inmunidad en la mucosa local de las vías respiratorias. Para obtener una respuesta inmunológica detectable debe administrarse una dosis suficiente por la vía apropiada. Este hecho se ilustra por la inmunización contra el tétanos: la calidad de toxina producida por las bacterias en la inyección inicial es insuficiente para desarrollar una reacción inmunológica detectable, haciendo que sean necesarias la inmunización y las inyecciones de refuerzo del toxoide tetánico.
Anticuerpos
Los anticuerpos son un grupo heterogéneo de proteínas séricas denominadas inmunoglobulinas, la mayor parte de las cuales emigran durante la electroforesis como gamaglobulinas. Se han identificado en el suero humano cinco clases principales de inmunoglobulinas: IgG, IgM, IgA, IgD e IgE. Estas clases se distinguen con base en sus propiedades electroforéticas y serológicas. Tal vez todas las inmunoglobulinas séricas son anticuerpos contra algún antígeno, pero en la actualidad es imposible demostrarlo.
Las inmunoglobulinas son muy heterogéneas; aún las moléculas de IgG que son específicas para sólo un antígeno constan de docenas de moléculas diferentes. Son secretadas por células B diferenciadas denominadas células plasmáticas. De acuerdo con la teoría de la selección clonal, una célula plasmática sólo producirá un anticuerpo específico. En algunas enfermedades como el mieloma múltiple, las proteínas de mieloma, que son el producto anormal de una clona única de células plasmáticas, son homogéneas y en la electroforesis se observan como una banda densa única. Ciertos anticuerpos producidos en respuesta a antígenos muy homogéneos, como el polisacárido del estreptococo, pueden ser homogéneos.
Los monómeros de inmunoglobulinas están formados de dos cadenas pesadas y dos cadenas ligeras idénticas [ver figura 3]. Cada cadena ligera está unida a una cadena pesada por puentes disulfuro (S-S) y las cadenas pesadas también están unidas entre sí por uno o más puentes S-S. Las secuencias de aminoácidos en las cadenas pesadas y ligeras están divididas en regiones que son constantes o variables [ver figura 4]; además, cada región variable contiene regiones qe son hipervariables [ver adelante, Estructura y secuencia de las inmunoglobulinas].
CLASIFICACION DE LAS INMUNOGLOBULINAS
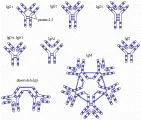
Existen cinco clases principales de inmunoglobulinas y cada clase está compuesta de una cadena pesada específica: la IgG tiene dos cadenas g; la IgM, dos cadenas µ; la IgA, dos cadenas a; la IgD, dos cadenas d y la IgE, dos cadenas e [ver tabla 1]. Así mismo, existen dos tipos de cadenas ligeras, k y l, las cuales pueden diferenciarse por medio de antígenos. La forma secretada de IgM es un pentámero compuesto de cinco monómeros [ver figura 5]; la IgM también puede existir como monómero unido a superficies. La IgA se encuentra en dos formas, como monómero o como dímero. Las inmunoglobulinas poliméricas, IgM e IgA, tienen una cadena J adicional (de unión), que ayuda en su polimeración.
IgG La IgG es la inmunoglobulina principal en el suero, donde se encuentra en forma de monómero. Es el principal anticuerpo que se eleva después de un reto antigénico. Aunque el feto no produce esta clase de inmunoglobulina, la IgG cruza con facilidad la placenta y, por tanto, cualquier cantidad de anticuerpos IgG que se encuentran en el recién nacido se habrá originado en la madre.
Los anticuerpos que cubren a los microrganismos haciéndolos más susceptibles a la fagocitosis (i.e., opsonización) son de la clase IgG. Los anticuerpos IgG también neutralizan toxinas, como la toxina diftérica, y pueden neutralizar virus.
La IgG tiene una vida media en sangre cercana a 23 días. Clínicamente se ha empleado con éxito para reconstituir la inmunidad de enfermos con agammaglobulinemia y para prevenir la enfermedad hemolítica del recién nacido. Las mujeres con grupo sanguíneo Rh negativo que tienen un feto con eritrocitos Rh1 positivos, pueden ser sensibilizadas durante el primer parto por los eritrocitos Rh1 positivos del feto. Entonces la madre producirá anticuerpos IgG anti-Rh1 que atravesarán la placenta durante embarazos subsecuentes; estos anticuerpos reaccionarán con los eritrocitos fetales, ocasionando la enfermedad hemolítica. En la actualidad esto puede prevenirse inyectando anticuerpos anti-Rh positivos, ricos en IgG (RhoGAM) a la madre Rh1-negativa, durante el parto o poco tiempo después de éste o de un aborto. Los anticuerpos parecen combinarse con cualquier eritrocito fetal Rh1-positivo presente, evitando que éstos inmunizen a la madre. Se desconoce el mecanismo exacto de esta inmunosupresión, pero tal vez sea un ejemplo del control por retroalimentación de la IgG en la producción de anticuerpos, es decir, se ha demostrado que la producción de IgG contra un antígeno es capaz de reducir aún más la producción de ese anticuerpo.
Existen cuatro subclases de IgG: IgG1, IgG2, IgG3 e IgG4, las cuales difieren en estructura y propiedades biológicas [ver tabla 2]. Por ejemplo, sólo la IgG1 y la IgG3 fijan al primer componente del complemento y se adhieren a los monocitos. Las funciones exactas de estas subclases son, en su mayor parte, desconocidas.
IgA La IgA es la inmunoglobulina predominante en las secreciones, donde suele encontrarse en forma de dímero y es liberada como tal por las células plasmáticas que se encuentran en la cercanía. La IgA está presente en saliva, lágrimas y calostro. También se encuentra en el aparato respiratorio y digestivo, en la vagina y en la próstata. El dímero se combina con una cadena polipeptídica, denominada pieza secretora, que es producida por las células epiteliales locales. De esta forma, es muy resistente a la digestión proteolítica. A diferencia de la IgA sérica, la IgA combinada con la pieza secretora se transporta en forma activa a través del epitelio mucoso por endocitosis [ver figura 6]. Se cree que la IgA juega un papel importante en la inmunidad local al neutralizar virus y al combinarse con virus y bacterias, impidiendo la adherencia de estos microrganismos a la superficie de las mucosas.
Aunque la IgA no fija complemento, se ha demostrado que es capaz de activar la vía alterna del mismo. Uno de los componentes del complemento generados por esta vía, el C3b, puede ayudar en la opsonización de las bacterias, y por lo tanto facilita su captación y eliminación por los fagocitos.
En su forma monomérica, la IgA constituye el 15 porciento de las inmunoglobulinas séricas. En suero, tiene una vida media de cinco a seis días. Existen dos clases de IgA, la IgA1 y la IgA2. Los niveles elevados de anticuerpos contra antígenos de la dieta que se encuentran en personas con deficiencia de IgA sugieren que esta inmunoglobulina en particular limita la absorción de esos antígenos.
IgM La IgM es un pentámero que se encuentra sobre todo en el suero, donde forma el 10 porciento de las inmunoglobulinas. Los estudios de microscopía electrónica revelan que tiene la forma de una estrella, pero cuando se encuentra unida a una bacteria sus sitios antigénicos están fijos a la superficie bacteriana lo cual hace que cambie su apariencia a una forma parecida a un cangrejo. Como un pentámero, la IgM es muy eficiente para fijar complemento. Molécula por molécula, es 20 veces más efectiva en aglutinar bacterias y eritrocitos y 1,000 veces más activa en las reacciones bactericidas que la IgG. Las isohemaglutininas, como la anti-A y la anti-B, son de la clase IgM, y la macroglobulinemia de Waldenström representa la producción monoclonal de IgM.
En la respuesta inmunológica, la IgM es la primera inmunoglobulina en elevarse tras el estímulo antigénico. El feto elaborará anticuerpos IgM contra ciertos microrganismos lo que puede ser de ayuda en el diagnóstico de la toxoplasmosis, la rubeola o la sífilis fetal; sin embargo, no todos los fetos infectados por esos microrganismos producen tales anticuerpos. Debido a que la respuesta de IgM contra antígenos es de corta duración, su presencia también puede ser de ayuda para establecer el diagnóstico de una infección en particular. Es posible determinar qué porcentaje de anticuerpos específicos en una muestra de suero consisten en IgM agregando una pequeña cantidad de mercaptoetanol, lo cual inhibe a esta clase de inmunoglobulinas y midiendo después lo que resta en la muestra.
Las células que producen IgM o sus precursores no se convierten en células de memoria, por lo que una segunda exposición a un antígeno no produce una mayor cantidad de anticuerpos IgM que el primer estímulo. La mayoría de las células B tienen IgM e IgD en su superficie [ver adelante, Receptores de inmunoglobulinas].
IgD La IgD, que es un monómero, existe en el suero en cantidades mínimas. Se descubrió primero como una proteína de mieloma que no pertenecía a ninguna de las clases conocidas. Se encuentra en concentraciones relativamente elevadas en la sangre del cordón umbilical. La mayoría de las células B de la sangre del cordón umbilical tienen IgD en su superficie y la mayoría de las células B del adulto tienen, en su superficie, tanto IgD como IgM. Así, se ha sugerido que la IgD podría ser un receptor celular temprano para antígenos; sin embargo, no existe evidencia de que la IgD sea precursora de la IgM. Se han encontrado células plasmáticas productoras de IgD en las amígdalas y adenoides, aunque su existencia es muy rara en otros tejidos linfoides. La función de la IgD se desconoce.
IgE La IgE se encuentra presente en cantidades mínimas en el suero, constituyendo sólo el 0.004 porciento de las inmunoglobulinas séricas. La IgE se une fuertemente a las células cebadas y a los basófilos; cuando estas células cubiertas con IgE interaccionan con antígenos específicos (alergenos) liberan potentes sustancias mediadoras de la hipersensibilidad inmediata, incluyendo a la histamina, la sustancia de reacción lenta de la anafilaxia (SRS-A) y un factor quimiotáctico eosinofílico. Por lo tanto, la IgE es el anticuerpo termolábil (que se conocía en el pasado como reagina) que desempeña una función principal en la hipersensibilidad inmediata, es decir, la reacción inmunológica que se presenta en la fiebre del heno, el asma extrínseco, la urticaria y la anafilaxia. Los niveles de IgE son mayores de lo normal en personas con atopia y los antígenos IgE específicos contra un alergeno en particular también están elevados, correlacionando con la susceptibilidad del individuo. En las alergias, los anticuerpos IgE específicos se detectan por medio de radioinmunoanálisis (RAST o prueba radioalergosorbente).
La IgE se une a las células cebadas o a los basófilos por medio de su porción Fc. El calor destruye la capacidad de la porción Fc de unirse a la célula. Se encuentran células plasmáticas productoras de IgE en las amígdalas, adenoides y la mucosa de los aparatos respiratorio y gastrointestinal. Receptores precisos para la IgE se localizan sobre la superficie de las células cebadas, las células B y las células T, los macrófagos y los eosinófilos.
Se desconoce la función exacta de la IgE. Ciertamente, las manifestaciones de la hipersensibilidad inmediata, como la fiebre del heno o el asma extrínseco, no parecen tener un propósito útil para el individuo afectado o para la especie en general. Por lo tanto, la observación de que los niveles de IgE se encuentran elevados en ocasiones en personas que viven en las regiones tropicales y en especial en aquéllas afectadas por parasitos helmintos, fue considerada por algunos inmunólogos como una posible indicación de la función de la IgE, es decir, que tiene un papel protector contra los parásitos. Los mediadores liberados podrían afectar a los parásitos, ya sea de manera directa o al producir incremento en la permeabilidad vascular y liberar factor quimiotáctico eosinofílico, lo cual podría causar el acúmulo de otros anticuerpos (IgG) y células necesarios para atacar al parásito. En este contexto es de interés el hecho de que los eosinófilos pueden mediar un daño dependiente de IgG contra esquistosomulas (la forma larvaria del parásito Schistosoma mansoni). Además, complejos inmunes de IgE específicos contra parásitos pueden inducir citotoxicidad mediada por macrófagos contra organimos esquistosómulas.
DIFERENCIAS ANTIGENICAS
Existen tres tipos de determinantes serológicos (antigénicos) en las inmunoglobulinas: isotípicos, alotípicos e idiotípicos.
Determinantes isotípicos Los determinantes isotípicos establecen la diferencia entre las regiones constantes de las diversas clases y subclases de cadenas ligeras; representan diferentes genes de la región constante [ver tabla 1]. Por ejemplo, en un individuo existen cuatro isotipos de la cadena pesada de IgG: g1, g2, g3 y g4, que representan las subclases IgG1, IgG2, IgG3 e IgG4, respectivamente. Hay un sólo isotipo de la cadena ligera k y un isotipo de la cadena ligera l.
Cerca del 60 porciento de las inmunoglobulinas están formadas de cadenas pesadas y ligeras k y 40 porciento consisten en cadenas pesada y ligeras l. La inmunización con ciertos antígenos induce la formación de anticuerpos con la misma distribución de determinantes k y l, que la encontrada en la población general de inmunoglobulinas; sin embargo, algunos antígenos provocan la formación de inmunoglobulinas compuestas sobre todo por cadenas k o l; del mismo modo, algunos antígenos inducen más la producción de una clase en particular de inmunoglobulinas que otros. Se desconoce qué mecanismos determinan la clase de inmunoglobulinas que se producen o qué factores controlan el tipo de cadena ligera que se unirá a las cadenas pesadas.
Determinantes alotípicos Los determinantes alotípicos distinguen entre las inmunoglobulinas de un isotipo en particular, representan alelos diferentes de los genes de las inmunoglobulinas y, por lo tanto, se determinan genéticamente de acuerdo con las leyes mendelianas de manera muy similar a la que determina los grupos sanguíneos ABO. Las moléculas de cadena pesadas g de un individuo pueden tener varios marcadores diferentes, mientras que no se conocen marcadores alotípicos para las cadenas pesadas µ, d y e o para las cadenas ligeras l. Las cadenas pesadas g poseen más de 20 marcadores alotípicos distintos, que se determinan en forma colectiva Gm. Además, las cadenas ligeras k contienen un grupo de marcadores alotípicos llamados Inv. Se han reconocido tres de estos marcadores: Inv 1, Inv 2 e Inv 3.
Determinantes idiotípicos Un idiotipo se define como un determinante antigénico único localizado en la región variable de un anticuerpo. Un idiotipo es la expresión antigénica de la región variable de un anticuerpo. Los determinantes idiotípicos distinguen a una inmunoglobulina de otra del mismo alotipo. Si un animal es inmunizado con una proteína de mieloma (i.e., una inmunoglobulina homogénea producida por células plasmáticas neoplásicas) el anticuerpo resultante tendrá anticuerpos dirigidos contra muchas partes de la molécula de mieloma. Cuando se extraen los anticuerpos de este antisuero que reaccionan con otras proteínas de mieloma del mismo isotipo y alotipo, el antisuero resultante seguirá siendo capaz de reaccionar con las proteínas inmunogénicas del mieloma pero no con otras. Pueden realizarse experimentos similares utilizando anticuerpos monoclonales. Los antisueros reaccionan con determinantes localizados en la porción variable de las cadenas ligeras y pesadas (ver adelante). En situaciones en las que se encuentra que la proteína de mieloma es un anticuerpo dirigido contra un hapteno específico, la agregación de tal hapteno evitará que la proteína de mieloma reaccione con el suero anti-idiotípico [ver figura 7]. Esto indica que los determinantes idiotípicos están conectados de manera íntima con el sitio de unión del antígeno en la inmunoglobulina. Los anticuerpos inducidos por un antígeno único tendrán diferentes especificidades idiotípicas, lo que ilustra una vez más la gran heterogeneidad de las inmunoglobulinas.
ESTRUCTURA Y SECUENCIA DE LAS INMUNOGLOBULINAS
Los estudios iniciales realizados con las proteínas de Bence Jones (dímeros de cadenas ligeras), con las proteínas de mieloma y con anticuerpos homogéneos purificados, produjeron un descubrimiento sorprendente: cuando se compararon anticuerpos de la misma clase, se encontró que una parte de cada cadena polipeptídica del anticuerpo era la misma, pero otra parte era muy variable. En la actualidad se sabe que la porción terminal carboxilo (C) de todas las cadenas ligeras k tiene una secuencia de aminoácidos idéntica, y por lo tanto a esta región se le denomina dominio constante o C, mientras que la porción terminal amino (N) tiene una secuencia de aminoácidos variable denominada secuencia V [ver figura 4]. Los primeros 110 aminácidos de la porción N-terminal de la cadena l ligera y de la cadena pesada también son variables, mientras que el 75 porciento restante de la cadena pesada es constante y consiste en un grupo de tres regiones homólogas. La semejanza entre las regiones constantes de las cadenas ligeras y pesadas indica que éstas pueden haber evolucionado a partir de un gen ancestral común.
Las regiones variables de las cadenas ligeras y pesadas pueden dividirse en varios subgrupos. Por ejemplo, con base en las diferencias en sus regiones variables, se distinguen cuatro subgrupos de cadenas ligeras k y seis subgrupos de cadenas ligeras l. Existen también tres subgrupos de regiones variables en las cadenas pesadas
Dentro de las regiones variables existen tres áreas que muestran gran variación [ver figura 8], y que se conocen como regiones hipervariables, correspondiendo al sitio de combinación del antígeno con el anticuerpo.
El análisis por rayos-X ha demostrado que las moléculas de las
inmunoglobulinas se forman de unidades globulares compactas unidas por
segmentos cortos de cadenas polipeptídicas más o menos lineales
[ver figura 4]. Las secuencias de los aminoácidos de todos los
dominios de las inmunoglobulinas comparten el mismo esquema plegable
básico, que consiste en siete cadenas b organizadas en dos hojas con
plegamiento b antiparalelo [ver figura 9]. La estructura básica,
denominada pliegue de Ig, se encuentra en todos los componentes de la familia
del supergen de las inmunoglobulinas, incluyendo los marcadores de superficie
de las células T, CD3-e, CD3-d, CD4 y CD8.
La IgG puede dividirse en tres fragmentos por la papaína, una enzima preteolítica [ver figura 10]. Dos de estos fragmentos son similares y se denominan Fab; el tercer fragmento se denomina Fc. La porción Fc es responsable de la actividad biológica de las diversas inmunoglobulinas; entre otras funciones, la porción Fc controla la capacidad de las inmunoglobulinas para unirse a las células, fijar complemento y atravesar la placenta. Puede observarse que la papaína actúa sobre el extremo amino terminal de los puentes S-S que mantienen unidas a las cadenas pesadas entre sí.
Otra enzima proteolítica, la pepsina, divide a la molécula de IgG más allá de los puentes S-S que unen a las cadenas pesadas, dejando un fragmento de gran tamaño, F(ab')2, el cual tiene la capacidad de unir y precipitar antígenos por su valencia doble y por su capacidad para formar una malla. En este procedimiento, la digestión enzimática divide a la fracción Fc en muchos fragmentos pequeños. La porción que es susceptible a la digestión por ambas enzimas se denomina región de unión o de bisagra, las enzimas proteolíticas pueden tener acceso a esta región porque contiene muchos residuos de prolina que evitan que la cadena peptídica se pliegue y forme una hélice.
Los anticuerpos catalíticos son un tipo novedoso de inmunoglobulina.6,7 Estos anticuerpos monoclonales se originan contra haptenos que recuerdan el estado de transición de una reacción enzimática, como la hidrólisis del éster carboxílico. Los anticuerpos se comportan como enzimas cuando se mezclan con las sustancias adecuadas.
SINTESIS DE ANTICUERPOS (IN VITRO E IN VIVO)
Cuando el antígeno se introduce por primera vez al organismo, se presenta una respuesta primaria que se caracteriza por una fase latente, que dura varios días, durante la cual no se detectan anticuerpos; en seguida se produce un aumento en la cantidad en suero de anticuerpos IgM, que suelen alcanzar un nivel máximo después de siete días. Al cabo de seis a siete días, también se detectan anticuerpos IgG. El pico de IgM empieza a disminuir antes de que se alcance el título máximo de IgG después de 10 a 14 días; a continuación, los títulos de anticuerpos disminuyen y muy poco antígeno se detecta cuatro a cinco semanas después de una dosis única de antígeno. En un segundo encuentro con el antígeno se producirá una respuesta secundaria (también denominada respuesta anamnésica o de refuerzo). Tanto los títulos de IgM como los de IgG se elevan de manera exponencial, sin la fase latente que se observa en la respuesta primaria [ver figura 11]. Mientras que el pico de IgM puede ser de la misma magnitud o ligeramente mayor que el que se presenta durante la respuesta primaria, la respuesta de la IgG es mucho mayor y dura más tiempo. Esta variación en la respuesta es una demostración apropiada de la memoria inmunológica, y se debe a la proliferación de células B antígeno específicas y de células T cooperadoras durante la respuesta primaria. Las características de las respuestas primaria y secundaria explican la necesidad de inyecciones de refuerzo en los programas de inmunización.
Maduración de la afinidad
Las propiedades de unión del anticuerpo cambian con el tiempo. A este proceso se le denomina maduración de la afinidad, e incluye el proceso de maduración somática y de selección. Después del estímulo primario, los anticuerpos tienen una afinidad cada vez mayor por el antígeno según va pasando el tiempo, y forman complejos antígeno-anticuerpo con una mayor estabilidad. Además, los anticuerpos se vuelven menos específicos y presentan más reacciones cruzadas con antígenos relacionados. En realidad, la disminución en la especificidad refleja el hecho de que las reacciones cruzadas, que previamente eran muy débiles para ser detectadas, se vuelven aparentes al tener el anticuerpo mayor afinidad por el antígeno.
Mecanismos de control
La IgG no se incrementa de manera indefinida, deben existir algunos mecanismos de control para detenerla. Si esto no ocurriera, las células proliferantes escaparían al control y saturarían al organismo con anticuerpos. De hecho, parecen existir varios mecanismos de control; primero, disminuye la cantidad de antígeno. Segundo, existen evidencias considerables de que la IgG específica contra antígeno suprime por sí misma la síntesis de IgG. Aunque puede suponerse en primer término que esta reacción se produce por el simple bloqueo del antígeno, esta explicación no es válida por completo debido a que la porción F(ab')2 de la IgG no suprime la síntesis de IgG de una manera tan eficaz como la molécula completa de IgG. Tercero, tal vez participe la regulación idiotípica [ver figura 7]. Por último, las células T supresoras también ejercen control sobre la producción de anticuerpos. De hecho, al encontrar títulos inexplicablemente elevados de IgE en seres humanos, debe considerarse la posibilidad de una falta de células T supresoras.
Competencia antigénica
Otro ejemplo del control de la produción de anticuerpos es el fenómeno de competencia antigénica. Si ciertos antígenos se administran de manera secuencial o combinados, un antígeno impedirá la producción de anticuerpos contra el segundo; por ejemplo, a menos que los toxoides tetánicos y diftérico sean administrados en la proporción correcta, un antígeno impedirá una respuesta inmunológica contra el otro. Así mismo, cuando se inyecta una inmunoglobulina (i.e., a un conejo) se presenta una elevación de anticuerpos mucho mayor contra la porción Fc que contra la porción Fab. Si se inyectan cantidades iguales de fragmentos Fab y Fc, entonces se producen anticuerpos contra los dos.
El análisis del fenómeno de la competencia antigénica sugiere que varios mecanismos se encuentran relacionados. Es probable que uno de éstos sea la producción, por las células T, de factores supresores no específicos. Puede suceder que el primer antígeno administrado o el antígeno dominante estimulen la producción de factores de las células T, los cuales eviten la respuesta contra el segundo antígeno. Este posible mecanismo de la competencia antigénica puede ser importante en la inmunosupresión ocasionada por virus. De manera adicional, en algunos casos existe un límite en el número de células B antígeno-específicas que pueden competir por los dos antígenos, lo cual es probablemente el mecanismo que limita a los anticuerpos contra la porción Fc de la molécula de la IgG cuando se inyecta la inmunoglobuliina completa.
Cambio en la producción
Como se mencionó antes, la producción de IgM va seguida por la producción de IgG. En la actualidad existe evidencia considerable de que las células productoras de IgM cambian hacia la producción de IgG [ver figura 12]; por ejemplo, la administración de anticuerpos dirigidos contra la cadena pesada de la IgM, la cadena µ, puede evitar la producción subsecuente tanto de IgM como de IgG, mientras que los anticuerpos contra la cadena pesada de la IgG, la cadena g, sólo evitan la síntesis de la IgG. Más aún, algunos pacientes tienen mielomas de las clases IgM e IgG y el análisis ha demostrado que estos mielomas comparten el mismo idiotipo. En otras palabras, las regiones variables de sus cadenas ligeras y pesadas son las mismas, pero las regiones constantes de sus cadenas pesadas difieren. Estas observaciones no sólo son de importancia para comprender la manera en que las células cambian de la producción de IgM a la producción de IgG, sino que también tienen implicaciones importantes para el control genético de la síntesis de inmunoglobulinas.
INMUNOGLOBULINAS COMO RECEPTORES
Receptores de células B
Además de secretarse, las inmunoglobulinas pueden expresarse en la superficie de las células B, en donde actúan como receptores de antígenos.8 Estas inmunoglobulinas de superficie de membrana (IgSm) difieren de las inmunoglobulinas secretadas en que tienen un dominio transmembrana y son monoméricas. La primera IgSm que expresa una célula B es IgM, en una fase más avanzada del desarrollo se expresa en forma conjunta IgD [ver adelante, Origen genético de la diversidad de los anticuerpos]. La IsSm no viaja a la superficie celular por sí misma, sino que el proceso requiere la formación de un complejo consistente en la Ig y dos cadenas polipeptídicas denominadas Iga e Igb en el retículo endoplásmico [ver figura 13a]. El receptor de la célula B resultante se une a un antígeno. La célula B procesa el antígeno y expresa un fragmento peptídico en una molécula del CPH de clase II en la superficie celular para presentarlo a una célula T cooperadora. La estimulación de la célula T activa a la célula B, haciendo que se diferencíe a una célula de memoria o plasmática, que produce anticuerpos secretorios específicos para el antígeno encontrado. Después de la diferenciación terminal no se expresan más Iga o Igb. La célula plasmática madura cesa de expresar IgSm, a pesar de que mantenga el ARN mensajero (ARNm) para ésta.
Receptores para Fc
Se expresan receptores para Fc en múltiples células, incluyendo células cebadas, macrófagos, eosinófilos y células tumorales. Los receptores para Fc están compuestos de una familia de moléculas y su función es fijar la porción Fc de las inmunoglobulinas a la célula. El receptor de Fc para la IgE (RFce) sirve como modelo para todos los receptores de este tipo [ver figura 13b].9 Consiste en tres cadenas polipeptídicas, designadas como a, b, y g. La cadena RFce a es el sitio de unión para la porción Fc de la IgE. La RFceb es una molécula transmembrana que conecta a RFcea con la RFceg, la cadena responsable de reclutar las moléculas encargadas de las señales de transducción.
ORIGEN GENETICO DE LA DIVERSIDAD DE LOS ANTICUERPOS
El aspecto más intrigante del control genético de la síntesis de inmunoglobulinas es la diversidad del producto: las células plasmáticas pueden producir anticuerpos que reaccionen con más de 109 sitios antigénicos diferentes. ¿Cómo puede el ADN codificar contra un número tan grande de antígenos, muchos de los cuáles existen (en la escala de la evolución) desde hace muy poco tiempo?
La respuesta a esta pregunta ha empezado a surgir de los trabajos realizados utilizando técnicas de ADN recombinante.10-12 En estos estudios, el ADN de células embrionarias o germinales (espermatozoides) se comparó con el ADN, el ARN nuclear y el ARN mensajero (ARNm) de plasmocitomas. Los resultados indican, por ejemplo, que el ADN embrionario de la cadena ligera k codifica más de 300 regiones variables (V), cinco regiones de unión (J) y una región constante (C). Las regiones V y J se encuentran separadas del segmento C por una porción de ADN. Por lo tanto, el ADN embrionario tiene la capacidad para formar al menos 1,500 combinaciones diferentes de V y J; en otras palabras, por lo menos 1,500 cadenas ligeras k diferentes.
La aparición de estas líneas celulares individuales es el resultado de la recombinación somática en el ADN y de la unión del ARN [ver figura 14]. Al diferenciarse las células embrionarias en células plasmáticas, delecciones en el ADN hacen que uno de los genes V, escogidos al azar, sea adyacente a uno de los genes J. Esta unidad V-J y el resto de las regiones J están separadas del gen C por una porción corta de ADN. En el siguiente paso, el ADN es trancrito al ARN nuclear y la secuencia queda lista para un segundo rearreglo.
Este evento se inicia cuando una enzima rompe el ARN nuclear para producir ARN mensajero (ARNm). En un proceso denominado unión del RNA, el segmento que separa a la región o unidad V-J de la región C es removido junto con los segmentos J sobrantes. El segmento V-J-C restante se traduce en una de las 1,500 o más proteínas de las cadenas ligeras k. En realidad el número es mayor por el hecho de que la unión de cualquier porción V con J puede afectar alguna entre una variedad de secuencias de bases apareadas en el sitio de recombinación.
La variabilidad en la cadena pesada también contribuye de manera importante a la especificidad de un anticuerpo. Esta variabilidad es consecuencia de un proceso que incluye la recombinación somática y la unión de ARN, similares al proceso que crea la variabilidad en las cadenas ligeras. El ADN embrionario lleva las instrucciones para la formación de varios cientos de genes V de cadena pesada, seis regiones J, 10 a 20 regiones D (diversidad) y nueve genes C (estos genes C codifican las clases de cadenas pesadas: IgM, IgD, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2 e IgE). La deleción de ADN, su transcripción al ARN nuclear y la unión del ARN producen la secuencia V-C-J-J final en el ARNm, que es codificada por los ribosomas a una proteína de cadena pesada.
La operación aritmética se encuentra ahora completa: puede calcularse que el mecanismo ADN-ARN de cadena pesada produce más de 18,000 variedades posibles (la especificidad del anticuerpo no varía con la clase del mismo, por lo que los nueve genes C no entran en el cálculo). La combinación de las más de 1,500 variedades de cadenas ligeras con las 18,000 variedades de cadenas pesadas podría producir más de 27 millones de anticuerpos diferentes con sitios de combinación para antígenos diferentes.
De esta manera, parecería que la diversidad de los anticuerpos en las cadenas ligeras k y en las cadenas pesadas puede ser producida por la recombinación somática de los genes de las líneas germinales. Sin embargo, esta no es la única base para la generación de diversas moléculas de anticuerpos. Los estudios realizados en la cadena ligera l indican un mecanismo adicional. Estas cadenas son muy similares entre sí y sólo muestran variación ocasional en su secuencia de nucleótidos. En vista de que sólo se ha encontrado un segmento Cl1 por genoma haploide, sólo las mutaciones en punto pueden explicar este hallazgo. Se han encontrado variantes somáticas semejantes en las cadenas ligeras k y en las cadenas pesadas. La frecuencia de mutación somática es relativamente elevada (un par de bases por cada 1,000 divisiones celulares).
Las secuencias cortas y muy conservadas de ADN que limitan los segmentos V, D y J son utilizadas por la célula como señales para el rearreglo de los genes de las inmunoglobulinas [ver figura 15].10,12 Estas secuencias de ADN consisten en siete pares de bases (heptámero) y de una secuencia de nueve pares de bases (nonámero) separadas por 12 ó 23 pares de nucleótidos al azar, que constituyen los nucleótidos de dispersión. Una vuelta de la hélice del ADN equivale aproximadamente a doce pares de bases; por lo tanto, las secuencias que contienen nucleótidos de dispersión de doce pares de bases se denominan señales de una vuelta. En forma análoga, las secuencias que contienen nucleótidos de dispersión de 23 pares de bases, equivalentes a dos vueltas de la hélice de ADN, se denominan señales de dos vueltas. La recombinación de los segmentos de los genes de las inmunoglobulinas parece depender del apareamiento de estas señales de recombinación complementaria; sin embargo, al parecer el apareamiento sólo ocurre entre las señales de una de dos vueltas (la regla de unión de una o dos vueltas). La distribución de los dos tipos de secuencias de recombinación garantiza que sólo puedan ocurrir ciertas recombinaciones de genes.
El mecanismo de rearreglo V-D-J es característico de las células
inmunocompetentes, y emplea el mecanismo general de reparación del ADN.
Dos de los genes incluidos en el rearreglo V-D-J en las células T y B
son los genes activadores de la recombinación rag1 y
rag2.13 La ruptura de estos genes causa un bloqueo en el
desarrollo de las células B antes de la transición de
células pro-B a pre-B.14 Del mismo modo, la alteración
del gen de la IgM de superficie, de la región J de la cadena pesada o de
la región J de la cadena ligera k causa un bloqueo parecido en el
desarrollo de las células B.
Los genes que codifican a las cadenas pesadas de IgM e IgD (los genes µ y d) desempeñan una función muy importante en la respuesta primaria, que ocurre como resultado de la primera exposición al antígeno. Las células B iniciales expresan sólo IgM sobre su superficie; más tarde en la ontogenia, se expresa la IgM en la membrana celular junto con el producto final del gen de IgD. La IgM aumenta durante la ontogenia, alcanzando concentraciones en las células plasmáticas completamente diferenciadas alrededor de 100 veces más altas que las concentraciones en las células pre-B. Cuando las células B se diferencían, la IgD unida a la membrana disminuye hasta una concentración aproximada del 10 porciento de la IgM unida a la membrana. Junto con estos cambios cuantitativos ocurre una mayor modulación del desarrollo de las formas secretadas y unidas a la membrana de cada cadena pesada. Las formas de membrana de IgM e IgD predominan en las etapas iniciales en vista de que constituyen receptores para antígenos específicos. Sin embargo, ocurre un cambio al final de la diferenciación que ocasiona la producción casi exclusiva de la forma secretada de IgM. Aunque la IgD es un isotipo abundante en la superficie celular, es secretada sólo en pequeñas cantidades.15,16
En el cambio de producción de IgM a IgG que constituye la respuesta de refuerzo, los genes de la región constante son separados del ADN de la célula plasmática antes de que ocurra la transcripción al ARN.17,18 Por ejemplo, si la célula cambia hacia la producción de IgG3, los genes para µ y d son suprimidos [ver figura 16]. Después de la transcripción, la unión del ARN produce un mensajero con la secuencia V-D-J-Cg3, la cual se traduce a la proteína.
HIBRIDOMAS DE LINFOCITOS
El desarrollo del hibridoma de linfocitos, productos de fusión celular, tiene implicaciones revolucionarias para la inmunología y la medicina clínica.19 Los hibridomas de linfocitos constituyen el medio por el cual pueden producirse títulos muy altos de anticuerpos puros, muy específicos, para fines experimentales. Con este recurso los inmunólogos han sido capaces de incrementar y profundizar el conocimiento de las funciones del sistema inmune. Por ejemplo, los anticuerpos monoclonales han ayudado a definir los productos genéticos del CPH y muchos marcadores de superficie de las células hematopoyéticas, y han sido importantes para estudiar las bases de la mutación somática en los genes de los anticuerpos. En el campo clínico, la producción de estos anticuerpos ha permitido realizar pruebas serológicas de mucha mejor calidad. Se han usado anticuerpos monoclonales para eliminar células T y células tumorales antes del trasplante de médula ósea, y también en receptores de trasplante para evitar el rechazo de riñón o hígado al disminuir el número de células T citotóxicas.
El hibridoma de linfocitos, tal y como fue desarrollado por Köhler y Milstein, es el producto de la fusión de una célula de mieloma de ratón y un linfocito obtenido del bazo de un ratón inmunizado con un antígeno específico. La fusión puede lograrse empleando una variedad de virus o polietilenglicol. El resultado es una célula única con dos núcleos. Después, la membrana nuclear se rompe y los cromosomas se dividen y se distribuyen al azar entre las dos células hijas, los hibridomas de linfocitos.
Estas manipulaciones se llevan a cabo en un medio selectivo. Las células de mieloma, las cuales son incapaces de crecer en este medio, debido a que carecen de las enzimas necesarias, mueren. Aunque las células esplénicas poseen esta enzima, también mueren, debido a que no son capaces de replicarse de manera indefinida; sin embargo, las células híbridas tienen tanto la enzima del linfocito como la capacidad de la célula de mieloma para sobrevivir de manera indefinida, de manera que viven y al hacerlo inmortalizan la capacidad de la célula esplénica para elaborar anticuerpos específicos.
Los híbridos pueden crecer y ser clonados. El medio de cultivo de las clonas se prueba en busca de anticuerpos contra el antígeno deseado, de manera que las clonas positivas pueden ser seleccionadas y cultivadas en masa, in vitro o in vivo. De cualquier manera, debido a que se originan de una clona única, las células productoras de anticuerpos elaboran un solo tipo de anticuerpo en grandes cantidades. En comparación, las técnicas clásicas producen sueros que contienen un amplio espectro de anticuerpos diferentes al deseado, aun cuando la titulación sea muy elevada.
Un ratón que tenga hibridomas de linfocitos producirá anticuerpos específicos (por ejemplo, contra un antígeno de superficie linfocitario), en una cantidad mayor que todos los anticuerpos contra este antígeno sintetizados por muchos investigadores en todo el mundo. Tal anticuerpo puede tener títulos por millones, un fenómeno sin precedentes; así mismo, debido a que el anticuerpo se dirige contra un antígeno único, puede ser empleado para aislar ese antígeno.
ANTICUERPOS MONOCLONALES HUMANOS
En la actualidad existen varios métodos para elaborar anticuerpos monoclonales humanos. El primero consiste en tomar el ADN complementario que codifica un anticuerpo monoclonal murino y remplazar en forma sistemática las secuencias murinas con secuencias humanas. El ADN que codifica para las regiones constantes de las cadenas pesadas y ligeras humanas remplaza las secuencias de ADN murinas de las regiones variables con humanas por medio de mutagénesis, dejando las regiones hipervariables intactas. Una desventaja de este método es que el marco de lectura de las regiones variables puede afectar la afinidad. Por lo tanto, las inmunoglobulinas humanizadas pueden requerir mutaciones adicionales para aumentar su afinidad.
Otro método de humanizar anticuerpos monoclonales murinos consiste en tomar un segmento grande de ADN humano que incluya varias de las regiones variables humanas y todas las regiones constantes humanas e introducir este segmento en el genoma murino. Este ratón transgénico se cruza con un ratón en el que exista ruptura dirigida de la región J de los genes de las cadenas pesadas de Ig y ligeras k.20 Puede entonces inyectarse cualquier antígeno al ratón transgénico, y producirá anticuerpos monoclonales humanizados. Debido a que el ratón transgénico contiene un número limitado de regiones variables humanas, la potencia de estos anticuerpos depende de la mutación somática natural y de la maduración de la afinidad que ocurra en el ratón.
Un tercer método de generar anticuerpos monoclonales humanos consiste en construir bibliotecas (inventarios) de regiones variables humanas en bacterias o en bacteriófagos. El anticuerpo puede expresarse en la superficie de la bacteria o el bacteriófago y seleccionarse por afinidad al antígeno de interés.
Los anticuerpos monoclonales son de gran utilidad para investigar los mecanismos de la respuesta inmune. Por ejemplo, se usan para aislar antígenos de la superficie celular y definir subgrupos de linfocitos o de otras células. Son un instrumento ideal para producir grandes cantidades de anticuerpos homogéneos para estudiar la estructura fina del sitio de combinación del anticuerpo con un antígeno específico. También se han usado los hibridomas de linfocitos para determinar la base genética de la generación de la diversidad en la formación de anticuerpos. Administrados in vivo, los anticuerpos monoclonales permiten investigar el papel de varios subtipos de linfocitos que regulan el sistema inmune, al eliminarlos o activarlos. También se emplean para neutralizar diversas citocinas y para definir su función en la inmunorregulación e inflamación.
Los anticuerpos monoclonales pueden ser muy útiles en otros campos además de la inmunología. Por ejemplo, los anticuerpos contra los receptores hormonales pueden ser capaces de unirse al receptor y modificar una respuesta endócrina.
Receptores de células T
A diferencia de los receptores de inmunoglobulinas de las células B, que pueden reconocer antígenos libres, los receptores de células T sólo reconocen antígenos combinados con antígenos autólogos del complejo principal de histocompatibilidad, por lo general sobre la superficie de los macrófagos. Las células T cooperadoras o CD4+ (así como las pocas células T citotóxicas CD4+), requieren moléculas clase II del CPH, y las células T citotóxicas CD8+ requieren moléculas clase I del CPH. Este fenómeno se conoce como restricción del CPH. La capacidad de las células T para reconocer estas moléculas del CPH propias se establece en el timo antes de que los linfocitos se expongan al antígeno .
Las moléculas que forman el receptor de la célula T y los genes que codifican estas moléculas se han aislado y se están definiendo mediante el uso de anticuerpos monoclonales que reconocen las clonas de células específicas (clonotípicas), hibridomas y linfomas de células T21-23 y con el empleo de tecnología de ADN recombinante.24-27 Existen datos convincentes de que el receptor de las células T está compuesto de seis polipéptidos distintos conocidos como complejo CD3-receptor de células T (RCT-CD3)28 [ver figura 17]. Del 85 al 95 porciento de los linfocitos de la sangre periférica tienen receptores RCT-ab y sólo el cinco al 15 porciento tienen RCT-gd. Parece que los receptores gd son expresados por un subgrupo de linfocitos derivados del timo diferentes a las células que expresan RCT-ab. La porción del complejo RCT-ab que reconoce al antígeno está compuesta de dos cadenas polipeptídicas glucosiladas, denominadas RCT-a (45-55 kd) y RCT-b (37-45 kd), que están unidas por puentes disulfuro, formando un heterodímero de 90 kd. Las cadenas polipeptídicas correspondientes en el complejo RCT-gd consisten en RCT-g (45 a 55 kd)29 y RCT-d (35 a 45 kd). El mapeo de péptidos y la identificación de la secuencia de los aminoácidos de los heterodímeros en clonas de linfocitos con diferentes especificidades antigénicas, muestran que cada una de las cadenas de RCT-a, RCT-b, RCT-g y RCT-d contienen porciones variables (V) y constantes (C) análogas a las observadas en las inmunoglobulinas. No resulta sorprendente la presencia de regiones variables en moléculas que deben reconocer diversos antígenos. Las células RCT-ab pueden funcionar como células cooperadoras o como células asesinas, pero la única función relacionada con las células RCT-gd es la citotoxicidad. Se ha demostrado que algunas células RCT-gd producen lesiones cutáneas en la lepra.30
Los heterodímeros de RCT-ab y RCT-gd están muy relacionados con las proteínas CD3: CD3-g (25 a 28 kd), CD3-d (20 kd), Cd3-e (20 kd) y CD3-s (16 kd) [ver figura 16]. Se ha utilizado un grupo de anticuerpos monoclonales, incluyendo OKT3, anti-Leu-4 y UCHT1, para identificar las proteínas CD3 (con mayor frecuencia (CD3-e) en todas las células periféricas de sangre periférica y en el 30 al 60 porciento de los timocitos. Todos los demás timocitos, excepto un pequeño porcentaje, expresan una baja concentración de CD3 y RCT en su superficie.
Al llegar al timo para convertirse en una célula T, el pretimocito expresa genes CD3 de inmediato, pero solo en el interior de la célula. Este timocito CD3-, RCT- en la superficie (CD3+ en el citoplasma) se desarrolla en un compartimiento del timo que no tiene moléculas de superficie CD4 y CD8, por lo que se denomina timocito CD4-, CD8- o doble negativo (DN). En un subcompartimiento de los timocitos DN ocurre rearreglo y expresión del locus RCT-b (determinado por los genes rag1 y rag2, además de otros genes desconocidos). Por el contrario, el rearreglo y expresión del RCT-a ocurre en una etapa más tardía del desarrollo, en un compartimiento en el que los timocitos expresan moléculas de superficie tanto CD4 como CD8 para volverse timocitos CD4+, CD8+ o doble positivas (DP). El complejo RCT-CD3 se expresa completo en la superficie celular solo hasta esta fase del desarrollo de los timocitos DP. Una vez expresado, el complejo RCT-CD3 es responsable del proceso de selección positiva o negativa. Para la selección positiva el complejo RCT-CD3 tiene que reaccionar con un fragmento peptídico unido a una molécula de clase I del CPH para producir una célula T CD8+ o con un fragmento peptídico unido a una molécula de clase II del CPH para producir una célula T CD4+, ambas conocidas como células sencillas positivas. Esta vía de desarrollo ha sido confirmada por mutaciones del RCT producidas en ratones en los que se han eliminado genes específicos [ver figura 18]. Los ratones en los que se eliminaron los genes rag1, rag2 u otros que gobiernan la expresión del locus RCT-b presentaron un bloqueo en el desarrollo de los timocitos a nivel del timocito DN.31 Por el contrario, la eliminación del gen del RCT-a causa bloqueo del timocito DP. La alteración de los genes CD3 provoca bloqueo parcial del desarrollo de los timocitos por la redundancia de los componentes del CD3.
La expresión de las proteínas CD3 y RCT sobre la superficie celular es dependiente en forma mutua: ninguno de los componentes se observa sobre la superficie de las células T sin el otro. Por ejemplo, las células mutantes que no producen transcritos de ARNm de la cadena RCT-b carecen por completo de las proteínas de superficie CD3 y RCT; la transferencia de una copia normal del gen de la RCT-b en estas células restablece la transcripción y produce un nivel normal de las proteínas CD3 y RCT sobre la superficie celular. Además, la incubación de células T con anticuerpos dirigidos contra el RCT o el CD3 (o la incubación de células T antígeno-específicas con péptidos antigénicos específicos) causa comodulación, esto es, la desaparición del RCT y el CD3 de la superficie celular y su aparición en cuerpos multivesicales que más tarde se fusionan con lisosomas. La comodulación se acompaña de la pérdida de funciones antígenoespecíficas. El RCT y el CD3 se vuelven a expresar alrededor de 49 horas después de extraer los anticuerpos moduladores.
Los datos estructurales y funcionales indican que las actividades del receptor de células T se distribuyen entre las subunidades del complejo RCT-CD3: los polipéptidos del RCT (a, b, g y d) unen al antígeno y a los productos del gen del CPH, y las proteínas CD3 traducen la señal de unión al citoplasma de la célula T, lo que produce la activación de las funciones de la célula T.
RECEPTORES DE CELULAS T Y GENES CD3
La organización de los genes que codifican las cadenas a, b, g, y d del receptor de las células T humanas [ver figura 19] es análoga a la de los genes de las cadenas pesadas de las inmunoglobulinas; existen segmentos variables, de diversidad y de unión, que están limitados por secuencias de reconocimiento que modulan la recombinación antígeno específica [ver antes, Anticuerpos].32,33 De este modo, la diversidad de los receptores de las células T se genera en gran parte por los mismos mecanismos utilizados por las células B en la producción de las inmunoglobulinas.34 De hecho, las células T y B utilizan la misma enzima de recombinación, o recombinasa.34
Las secuencias genómicas que codifican la cadena RCT-b contienen dos genes de la región constante muy similares, Cb1 y Cb2, cada uno de los cuales se asocia con un grupo de seis o siete secuencias J y una secuencia D única [ver figura 19a]. Existen al menos 70 genes Vb que se relacionan con dos genes Cb. El rearreglo de los segmentos del gen de la cadena b permite la producción de aproximadamente 3,600 cadenas b diferentes.
Los genes del receptor a de las células T están distribuidas en forma diferente [ver figura 19b]. Un sólo gen Ca está precedido por una cadena muy grande de ADN compuesta de por lo menos 50 secuencias J distintas. No se ha demostrado de manera directa la presencia de un segmento del gen Da. Algunas secuencias Va se organizan como familias de genes relacionados. El rearreglo de los segmentos del gen de la cadena a puede representar cerca de 2,500 polipétidos diferentes. Por lo tanto, se pueden formar 9 X 106 RCTab. Aunque no se han detectado mutaciones somáticas en los receptores de células T, la recombinación imprecisa contribuye a la variabilidad de las cadenas RCT-a y RCT-b. Por consiguiente, la cantidad de RCT-ab puede ser considerablemente mayor de 9 X 106.
Los genes de cadenas RCR-b y RCT-g se han localizado en el cromosoma 7 y los genes de las cadenas RCT-a y RCT-d en el cromosoma 14, mediante el uso de híbridos de células somáticas de roedores y humanos y por técnicas de hibridación in situ [ver tabla 3]. Los genes para RCT-b y RCT-g se han localizado en brazos opuestos del cromosoma 7. Estos estudios de mapeo cromosómico indican que los genes del receptor de células T no están ligados al complejo principal de histocompatibilidad. Aunque existe cierta superposición en la localización cromosómica de los genes del receptor de células T y de las inmunoglobulinas, no hay consistencia entre las especies: los genes de la RCT-a y de las cadenas pesadas de las inmunoglobulinas se mapean en el cromosoma 14 en humanos pero se encuentran en cromosomas separados en el ratón, y los genes de las cadenas k de las inmunoglobulinas y de la RCT-b se localizan en el cromosoma 6 en el ratón pero no están relacionados en el genoma humano [ver tabla 3].
Los genes CD3-g, CD3-d CD3-e y CD3-s se transcriben en todas las células T, pero no se rearreglan. El CD3-g, CD3-d y CD3-e son codificados en la porción distal del brazo largo del cromosoma 11 humano (11q23-11qter); el CD3-s se localiza en el cromosoma 1 [ver tabla 3]. Los genes CD3-g, CD3-d y CD3-e se agrupan en una zona del cromosoma 11 con una extensión de cerca de 50,000 pares de bases. Su expresion específica en las células T es regulada por lo menos por dos factores potenciadores localizados justo hacia adelante de los genes CD3-d y CD3-e.35,36 Se considera que los genes CD3 desempeñan una función crucial en el inicio de la diferenciación intratímica. Estos genes de las cadenas de CD3 no están relacionados con los genes del CPH, del RCT o de las inmunoglobulinas, ni en humanos ni en ratones.
Respuesta inmune humana en modelos animales
Los ratones transgénicos son muy útiles en el estudio de la expresión del gen del receptor de las células T debido a un fenómeno conocido como exclusión alélica, que también ocurre en los genes de las inmunoglobulinas. La exclusión alélica se refiere al hecho de que una vez que ocurre el rearreglo del gen del RCT en un cromosoma, se inhibe el rearreglo subsecuente en el otro cromosoma. Por consiguiente, cuando se introduce un gen RCT-a o RCT-b rearreglado exógenamente en las células germinales de un ratón, se inhibe de manera notable el rearreglo endógeno de los genes RCT-a o RCT-b. Los estudios realizados en ratones transgénicos han demostrado que los mecanismos para la selección de autoantígenos (específicamente las moléculas de las clases I y II del CPH) desempeñan una función importante en la diferenciación final de las células T. Por ejemplo, en una serie de experimentos se aislaron los genes RCT-a y RCT-b de la clase I del CPH de una clona de células T citotóxicas CD4-, CD8+ específica para el antígeno H-Y masculino.37,38 Estos genes RCT-a y RCT-b antígeno-específicos rearreglados, fueron inyectados enseguida en los óvulos fertilizados murinos; la progenie que incorporó en su genoma el ADN inyectado (i.e., ratones transgénicos) fue utilizada para realizar más estudios.
El análisis de los timocitos y de las células T de ratones transgénicos demostró que la interacción específica entre el receptor de las células T sobre la superficie celular de timocitos inmaduros y los antígenos tímicos de la clase I del CPH determinaba las etapas finales de la diferenciación. Al parecer, las células T autorreactivas son eliminadas por selección negativa, ya que en los ratones varones, las células T autoespecíficas (i.e., específicas para el antígeno H-Y y restringidas por moléculas específicas de la clase I del CPH) son suprimidas en el timo. Varios estudios realizados en ratas transgénicas (sin el antígeno H-Y) aportaron datos acerca del reconocimiento intratímico de los antígenos de la clase I del CPH y demostraron que la selección positiva también afectaba la diferenciación de las células T inmaduras. En primer lugar, la frecuencia relativa de los timocitos CD4-, CD8+ fue mayor en los ratones transgénicos que en la especie del ratón original o en otros ratones que portaban alelos clase I del CPH diferentes. En segundo lugar, sólo las células T CD4-, CD8+ expresan altas concentraciones de cadenas polipeptídicas transgénicas RCT-a y RCT-b sobre su superficie celular. En tercer lugar, en los experimentos realizados para reconstruir la población celular de timos con células tronco de ratones transgénicos RCT-ab, ratones transgénicos RCT-b, y ratones normales, sólo las células madre transgénicas RCT-ab se asociaron con una proporción elevada de células CD4-, CD8+. Por lo tanto, parece que la interacción específica entre el receptor de las células T sobre los timocitos inmaduros CD4-, CD8+ y el antígeno de la clase I del CPH, provoca la diferenciación de estos timocitos en células T maduras CD4, CD8+. Otros experimentos indican que el reconocimiento de los antígenos de la clase II del CPH controla la diferenciación de células CD4-, CD8+ en células T maduras CD4-, CD8+. Por consiguiente, los modelos experimentales con ratones transgénicos apoyan la hipótesis de que los mecanismos de selección positiva y negativa intervienen en la diferenciación de timocitos humanos inmaduros en células T CD4+ y CD8+ con capacidad funcional.
El ratón "humano" con inmunodeficiencia combinada severa (ratón con IDCS-hu) es un modelo murino experimental utilizando para el análisis de la diferenciación y de la función hematolinfoide humana, pero también constituye una forma más directa para estudiar el sistema inmune humano que el modelo con ratones transgénicos.39-41 El ratón con IDCS tiene una deficiencia muy importante de linfocitos; las células de la población B o T no pueden detectarse por medio de estudios serológicos o funcionales. Al injertar células hematopoyéticas hepáticas, tejido del timo y de los ganglios linfáticos de fetos humanos al ratón con IDCS, se observó diferenciación de células humanas T y B maduras. Se ha detectado un aumento temporal de células T CD4+ y CD8+ humanas, además de IgG humana, en la circulación periférica de estos ratones con IDCS-hu. Las células T CD4+ humanas del ratón con IDCS-hu pueden ser infectadas por el virus del SIDA.42 El modelo de la IDCS-hu tendrá múltiples aplicaciones en el estudio de la respuesta inmune y de las enfermedades inmunológicas humanas.
Bibliografía
DR. COX THERHOST, PH.D.
DR. JOHN DAVID
Antígenos
Un antígeno es cualquier sustancia capaz de generar una respuesta inmune, esto es, de reaccionar con células T y células B para inducir la formación de anticuerpos y linfocitos sensibilizados, que reaccionan con esos anticuerpos y células una vez que son formados. Los primeros antígenos estudiados fueron diversas proteínas extrañas y de microrganismos, y sigue siendo válido que las proteínas son antigénicas en forma casi universal. Las bases de la inmunogenicidad de las proteínas se desconocen, pero es probable que se relacionen con su configuración única y estable. En la actualidad se sabe que en condiciones apropiadas un amplio rango de moléculas pueden inducir una respuesta inmune y que en situaciones anormales el organismo incluso desarrollará una respuesta inmune contra sus propios antígenos.
Las reglas que gobiernan la antigenicidad proteica se están estudiando con el uso de péptidos sintéticos seleccionados por la posición que ocupan en la estructura tridimensional de las proteínas [ver figura 1]. Casi todos los anticuerpos que aparecen en forma natural dirigidos contra proteínas son específicos de sitios determinados en la superficie de las moléculas proteicas.
|
|

Aunque la mayor parte de los antígenos son macromoléculas, algunas moléculas pequeñas también son antigénicas. Se ha demostrado que la hormona polipeptídica angiotensina amida, un octapéptido, y la arginina-vasopresina, un nonapéptido, pueden actuar como antígenos completos cuando se absorben en partículas de carbón. Así mismo, ciertos antibióticos conjugados con dinitrofenol (DNF), como la gramicidina-DNF y la bacitracina-DNF, actúan como antígenos al igual que el DNF(Lys), dinitrofenol conjugado con un polipéptido que contiene siete lisinas. Además, el ácido arsalínico azo- L-tirosina inducirá hipersensibilidad tardía sin formación de anticuerpos. Estas pequeñas moléculas antigénicas no contienen grupos reactivos, y por lo tanto, parecen ser antigénicas sin unirse a macromoléculas.
Se ha aprendido mucho acerca de los requerimientos de la inmunogenicidad a través del uso de polipéptidos sintéticos específicos. Por ejemplo, la agregación de aminoácidos aromáticos (como la tirosina), convertirá polipéptidos poco antigénicos en moléculas más inmunógenas aun cuando la especificidad del anticuerpo obtenido no esté dirigida contra este aminoácido. El uso de antígenos sintéticos permitió descubrir por primera vez que la respuesta inmune se encuentra bajo control genético; este descubrimiento evolucionó a partir de la observación de que algunas cepas de animales respondían a un polímero sintético, mientras que otras cepas no lo hacían.
Los antígenos no sólo son reconocidos por anticuerpos sino también a través de receptores de células T (RCT) antígeno-específicos, de los que existen dos tipos diferentes, denominados RCT-ab y RCT-gd. Los diversos tipos de células T tienen mecanismos diferentes para reconocer los antígenos. Por ejemplo, los RCT-ab reconocen antígenos que han sido procesados por células presentadoras de antígenos (CPA) para convertirse en fragmentos peptídicos unidos a moléculas del complejo principal de histocompatibilidad (CPH) de clase I o clase II en la superficie de las CPA. En contraste, los RCT-gd parecen no requerir la presentación de antígeno por el CPH. Las células T cooperadoras solo reconocen fragmentos peptídicos unidos a moléculas del CPH de clase II.1 Las células T citotóxicas reconocen antígenos procesados virales presentados tanto en moléculas de clase I como II del CPH sobre la superficie de las células humanas.2 En general, los receptores de las células T y las inmunoglobulinas reconocen diferentes determinantes antigénicos [ver adelante, Receptores de Células T]. Recientemente se ha demostrado que los lípidos puros de micobacterias pueden presentarse como antígenos al RCT-ab por las moléculas CD1 en lugar de las moléculas clase I o II del CPH.3
HAPTENOS
Existen algunas moléculas pequeñas que por sí solas no son antigénicas pero pueden reaccionar con anticuerpos; estas moléculas se denominan haptenos. Antes que ocurra la inducción de anticuerpos específicos por el hapteno, éste debe combinarse con una sustancia inmunógena, a menudo una proteína o un polipéptido sintético, denominada portador. Por ejemplo, si el hapteno fosforilcolina se combina con gamaglobulina bovina y el conjugado restante se inyecta a un animal, se producirán anticuerpos con predominio específico para el grupo fosforilcolina [ver figura 2]. Casi cualquier molécula que no sea antigénica por sí sola (diversos péptidos, esteroides, nucleótidos, lípidos y hormonas) pueden combinarse con un transportador inmunogénico y, como parte de un conjugado, inducir la formación de anticuerpos.
|
| Figura 2 |
| Sitio de unión anticuerpo-hapteno |
Esta respuesta de inducción es la base de potentes instrumentos de investigacipen el estudio de la respuesta inmune. Así mismo, es la base de muchos radioinmunoanálisis que se han convertido en auxiliares de diagnóstico útiles en medicina clínica. Además, la mayor parte de las reacciones por fármacos se producen por el hecho de que el medicamento agresor actúa como hapteno y se une en forma covalente con una proteína del propio individuo; tal vez el ejemplo más notable sea la unión covalente de la penicilina con proteínas séricas, que causa la reacción a la penicilina. La sensibilidad por contacto, incluyendo la inducida por metales, hiedra venenosa, roble venenoso, zumaque venenoso y detergentes, también es consecuencia de una molécula agresora que actúa como hapteno, que suele unirse a las proteínas de la piel.
Los anticuerpos contra haptenos de ácidos nucleicos pueden elaborarse en el laboratorio, pero esto ocurre rara vez in vitro. En pacientes con lupus eritematoso generalizado, enfermedad de Crohn o esclerosis lateral aminotrófica se producen anticuerpos antiADN y antiARN.
ADYUVANTES
Los adyuvantes son sustancias capaces de aumentar la inmunogenicidad de los antígenos. Muchos productos microbianos se han empleado como adyuvantes,4 incluyendo sustancias de Mycobacterium tuberculosis, bacilo de Calmette-Guérin (BCG), Corynebacterium parvum, Brucella abortus y Bordetella pertussis, así como toxoides de Vibrio cholerae y Clostridium tetani. Se han derivado adyuvantes del virus vaccinia y de otros poxvirus, BCG y Salmonella, cada uno transfectado con genes para un antígeno de interés. Otros son derivados de lipopolisacáridos, como el lípido monofosforil A. El adyuvante completo de Freund, que consiste en micobacterias muertas en aceite emulsificado con un antígeno en solución acuosa, se usa en la actualidad con poca frecuencia porque en general causa una intensa reacción inflamatoria local. Otros adyuvantes incluyen extractos de jabón del árbol de tejo (Quillaja saponaria), polímeros (v.gr., inulina), péptidos complejos y diversas citocinas. Por ejemplo, la interleucina-12 (IL-12) parece actuar como un promotor intenso de las reacciones inmunológicas mediadas por células en los ratones.5 La sal de aluminio alumbre también está autorizada para usarse como adyuvante en humanos.
Los liposomas también pueden actuar como adyuvantes. Las proteínas de membrana, como las proteínas de la pared viral insertadas en liposomas, son más antigénicas que aquellas solubilizadas en detergentes. En algunos casos se ha demostrado que los liposomas aumentan la respuesta inmune contra antígenos pequeños, lo cual indica que los liposomas pueden utilizarse en la elaboración de vacunas. En la práctica clínica, los adyuvantes se utilizan como complementos de algunas inmunizaciones, como la del tétanos, y son potencialmente importantes en el tratamiento antiparasitario y antitumoral.
RESPUESTA A ANTIGENOS
La respuesta del organismo a la exposición a antígenos depende de la vía de administración y de la dosis. En términos generales, los antígenos que entran al organismo por vía intravenosa viajan con rapidez al hígado, y la producción de anticuerpos es rápida. Sin embargo, la vía I.V. también facilita la inmunosupresión o la tolerancia, en especial cuando el antígeno es soluble y no agregado. Los antígenos que se administran por vía subcutánea o intradérmica suelen localizarse inicialmente en los ganglios linfáticos que drenan el sitio de la inyección, lo que favorece la inducción de inmunidad o la hipersensibilidad modulada por células. Los antígenos inhalados también favorecen la sensibilización local y el desarrollo de hipersensibilidad inmediata e inmunidad en la mucosa local de las vías respiratorias. Para obtener una respuesta inmunológica detectable debe administrarse una dosis suficiente por la vía apropiada. Este hecho se ilustra por la inmunización contra el tétanos: la calidad de toxina producida por las bacterias en la inyección inicial es insuficiente para desarrollar una reacción inmunológica detectable, haciendo que sean necesarias la inmunización y las inyecciones de refuerzo del toxoide tetánico.
Anticuerpos
Los anticuerpos son un grupo heterogéneo de proteínas séricas denominadas inmunoglobulinas, la mayor parte de las cuales emigran durante la electroforesis como gamaglobulinas. Se han identificado en el suero humano cinco clases principales de inmunoglobulinas: IgG, IgM, IgA, IgD e IgE. Estas clases se distinguen con base en sus propiedades electroforéticas y serológicas. Tal vez todas las inmunoglobulinas séricas son anticuerpos contra algún antígeno, pero en la actualidad es imposible demostrarlo.
Las inmunoglobulinas son muy heterogéneas; aún las moléculas de IgG que son específicas para sólo un antígeno constan de docenas de moléculas diferentes. Son secretadas por células B diferenciadas denominadas células plasmáticas. De acuerdo con la teoría de la selección clonal, una célula plasmática sólo producirá un anticuerpo específico. En algunas enfermedades como el mieloma múltiple, las proteínas de mieloma, que son el producto anormal de una clona única de células plasmáticas, son homogéneas y en la electroforesis se observan como una banda densa única. Ciertos anticuerpos producidos en respuesta a antígenos muy homogéneos, como el polisacárido del estreptococo, pueden ser homogéneos.
Los monómeros de inmunoglobulinas están formados de dos cadenas pesadas y dos cadenas ligeras idénticas [ver figura 3]. Cada cadena ligera está unida a una cadena pesada por puentes disulfuro (S-S) y las cadenas pesadas también están unidas entre sí por uno o más puentes S-S. Las secuencias de aminoácidos en las cadenas pesadas y ligeras están divididas en regiones que son constantes o variables [ver figura 4]; además, cada región variable contiene regiones qe son hipervariables [ver adelante, Estructura y secuencia de las inmunoglobulinas].
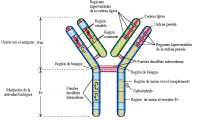
|
| Figura 3 |
| Estructura de la inmunoglobulina |
|
|
Existen cinco clases principales de inmunoglobulinas y cada clase está compuesta de una cadena pesada específica: la IgG tiene dos cadenas g; la IgM, dos cadenas µ; la IgA, dos cadenas a; la IgD, dos cadenas d y la IgE, dos cadenas e [ver tabla 1]. Así mismo, existen dos tipos de cadenas ligeras, k y l, las cuales pueden diferenciarse por medio de antígenos. La forma secretada de IgM es un pentámero compuesto de cinco monómeros [ver figura 5]; la IgM también puede existir como monómero unido a superficies. La IgA se encuentra en dos formas, como monómero o como dímero. Las inmunoglobulinas poliméricas, IgM e IgA, tienen una cadena J adicional (de unión), que ayuda en su polimeración.
|
| Figura 5 |
| Polímeros de inmunoglobulinas |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
IgG La IgG es la inmunoglobulina principal en el suero, donde se encuentra en forma de monómero. Es el principal anticuerpo que se eleva después de un reto antigénico. Aunque el feto no produce esta clase de inmunoglobulina, la IgG cruza con facilidad la placenta y, por tanto, cualquier cantidad de anticuerpos IgG que se encuentran en el recién nacido se habrá originado en la madre.
Los anticuerpos que cubren a los microrganismos haciéndolos más susceptibles a la fagocitosis (i.e., opsonización) son de la clase IgG. Los anticuerpos IgG también neutralizan toxinas, como la toxina diftérica, y pueden neutralizar virus.
La IgG tiene una vida media en sangre cercana a 23 días. Clínicamente se ha empleado con éxito para reconstituir la inmunidad de enfermos con agammaglobulinemia y para prevenir la enfermedad hemolítica del recién nacido. Las mujeres con grupo sanguíneo Rh negativo que tienen un feto con eritrocitos Rh1 positivos, pueden ser sensibilizadas durante el primer parto por los eritrocitos Rh1 positivos del feto. Entonces la madre producirá anticuerpos IgG anti-Rh1 que atravesarán la placenta durante embarazos subsecuentes; estos anticuerpos reaccionarán con los eritrocitos fetales, ocasionando la enfermedad hemolítica. En la actualidad esto puede prevenirse inyectando anticuerpos anti-Rh positivos, ricos en IgG (RhoGAM) a la madre Rh1-negativa, durante el parto o poco tiempo después de éste o de un aborto. Los anticuerpos parecen combinarse con cualquier eritrocito fetal Rh1-positivo presente, evitando que éstos inmunizen a la madre. Se desconoce el mecanismo exacto de esta inmunosupresión, pero tal vez sea un ejemplo del control por retroalimentación de la IgG en la producción de anticuerpos, es decir, se ha demostrado que la producción de IgG contra un antígeno es capaz de reducir aún más la producción de ese anticuerpo.
Existen cuatro subclases de IgG: IgG1, IgG2, IgG3 e IgG4, las cuales difieren en estructura y propiedades biológicas [ver tabla 2]. Por ejemplo, sólo la IgG1 y la IgG3 fijan al primer componente del complemento y se adhieren a los monocitos. Las funciones exactas de estas subclases son, en su mayor parte, desconocidas.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
* Células asesinas con citotoxicidad dependiente de anticuerpos. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
IgA La IgA es la inmunoglobulina predominante en las secreciones, donde suele encontrarse en forma de dímero y es liberada como tal por las células plasmáticas que se encuentran en la cercanía. La IgA está presente en saliva, lágrimas y calostro. También se encuentra en el aparato respiratorio y digestivo, en la vagina y en la próstata. El dímero se combina con una cadena polipeptídica, denominada pieza secretora, que es producida por las células epiteliales locales. De esta forma, es muy resistente a la digestión proteolítica. A diferencia de la IgA sérica, la IgA combinada con la pieza secretora se transporta en forma activa a través del epitelio mucoso por endocitosis [ver figura 6]. Se cree que la IgA juega un papel importante en la inmunidad local al neutralizar virus y al combinarse con virus y bacterias, impidiendo la adherencia de estos microrganismos a la superficie de las mucosas.
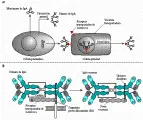
|
| Figura 6 |
| Secreción de IgA |
Aunque la IgA no fija complemento, se ha demostrado que es capaz de activar la vía alterna del mismo. Uno de los componentes del complemento generados por esta vía, el C3b, puede ayudar en la opsonización de las bacterias, y por lo tanto facilita su captación y eliminación por los fagocitos.
En su forma monomérica, la IgA constituye el 15 porciento de las inmunoglobulinas séricas. En suero, tiene una vida media de cinco a seis días. Existen dos clases de IgA, la IgA1 y la IgA2. Los niveles elevados de anticuerpos contra antígenos de la dieta que se encuentran en personas con deficiencia de IgA sugieren que esta inmunoglobulina en particular limita la absorción de esos antígenos.
IgM La IgM es un pentámero que se encuentra sobre todo en el suero, donde forma el 10 porciento de las inmunoglobulinas. Los estudios de microscopía electrónica revelan que tiene la forma de una estrella, pero cuando se encuentra unida a una bacteria sus sitios antigénicos están fijos a la superficie bacteriana lo cual hace que cambie su apariencia a una forma parecida a un cangrejo. Como un pentámero, la IgM es muy eficiente para fijar complemento. Molécula por molécula, es 20 veces más efectiva en aglutinar bacterias y eritrocitos y 1,000 veces más activa en las reacciones bactericidas que la IgG. Las isohemaglutininas, como la anti-A y la anti-B, son de la clase IgM, y la macroglobulinemia de Waldenström representa la producción monoclonal de IgM.
En la respuesta inmunológica, la IgM es la primera inmunoglobulina en elevarse tras el estímulo antigénico. El feto elaborará anticuerpos IgM contra ciertos microrganismos lo que puede ser de ayuda en el diagnóstico de la toxoplasmosis, la rubeola o la sífilis fetal; sin embargo, no todos los fetos infectados por esos microrganismos producen tales anticuerpos. Debido a que la respuesta de IgM contra antígenos es de corta duración, su presencia también puede ser de ayuda para establecer el diagnóstico de una infección en particular. Es posible determinar qué porcentaje de anticuerpos específicos en una muestra de suero consisten en IgM agregando una pequeña cantidad de mercaptoetanol, lo cual inhibe a esta clase de inmunoglobulinas y midiendo después lo que resta en la muestra.
Las células que producen IgM o sus precursores no se convierten en células de memoria, por lo que una segunda exposición a un antígeno no produce una mayor cantidad de anticuerpos IgM que el primer estímulo. La mayoría de las células B tienen IgM e IgD en su superficie [ver adelante, Receptores de inmunoglobulinas].
IgD La IgD, que es un monómero, existe en el suero en cantidades mínimas. Se descubrió primero como una proteína de mieloma que no pertenecía a ninguna de las clases conocidas. Se encuentra en concentraciones relativamente elevadas en la sangre del cordón umbilical. La mayoría de las células B de la sangre del cordón umbilical tienen IgD en su superficie y la mayoría de las células B del adulto tienen, en su superficie, tanto IgD como IgM. Así, se ha sugerido que la IgD podría ser un receptor celular temprano para antígenos; sin embargo, no existe evidencia de que la IgD sea precursora de la IgM. Se han encontrado células plasmáticas productoras de IgD en las amígdalas y adenoides, aunque su existencia es muy rara en otros tejidos linfoides. La función de la IgD se desconoce.
IgE La IgE se encuentra presente en cantidades mínimas en el suero, constituyendo sólo el 0.004 porciento de las inmunoglobulinas séricas. La IgE se une fuertemente a las células cebadas y a los basófilos; cuando estas células cubiertas con IgE interaccionan con antígenos específicos (alergenos) liberan potentes sustancias mediadoras de la hipersensibilidad inmediata, incluyendo a la histamina, la sustancia de reacción lenta de la anafilaxia (SRS-A) y un factor quimiotáctico eosinofílico. Por lo tanto, la IgE es el anticuerpo termolábil (que se conocía en el pasado como reagina) que desempeña una función principal en la hipersensibilidad inmediata, es decir, la reacción inmunológica que se presenta en la fiebre del heno, el asma extrínseco, la urticaria y la anafilaxia. Los niveles de IgE son mayores de lo normal en personas con atopia y los antígenos IgE específicos contra un alergeno en particular también están elevados, correlacionando con la susceptibilidad del individuo. En las alergias, los anticuerpos IgE específicos se detectan por medio de radioinmunoanálisis (RAST o prueba radioalergosorbente).
La IgE se une a las células cebadas o a los basófilos por medio de su porción Fc. El calor destruye la capacidad de la porción Fc de unirse a la célula. Se encuentran células plasmáticas productoras de IgE en las amígdalas, adenoides y la mucosa de los aparatos respiratorio y gastrointestinal. Receptores precisos para la IgE se localizan sobre la superficie de las células cebadas, las células B y las células T, los macrófagos y los eosinófilos.
Se desconoce la función exacta de la IgE. Ciertamente, las manifestaciones de la hipersensibilidad inmediata, como la fiebre del heno o el asma extrínseco, no parecen tener un propósito útil para el individuo afectado o para la especie en general. Por lo tanto, la observación de que los niveles de IgE se encuentran elevados en ocasiones en personas que viven en las regiones tropicales y en especial en aquéllas afectadas por parasitos helmintos, fue considerada por algunos inmunólogos como una posible indicación de la función de la IgE, es decir, que tiene un papel protector contra los parásitos. Los mediadores liberados podrían afectar a los parásitos, ya sea de manera directa o al producir incremento en la permeabilidad vascular y liberar factor quimiotáctico eosinofílico, lo cual podría causar el acúmulo de otros anticuerpos (IgG) y células necesarios para atacar al parásito. En este contexto es de interés el hecho de que los eosinófilos pueden mediar un daño dependiente de IgG contra esquistosomulas (la forma larvaria del parásito Schistosoma mansoni). Además, complejos inmunes de IgE específicos contra parásitos pueden inducir citotoxicidad mediada por macrófagos contra organimos esquistosómulas.
DIFERENCIAS ANTIGENICAS
Existen tres tipos de determinantes serológicos (antigénicos) en las inmunoglobulinas: isotípicos, alotípicos e idiotípicos.
Determinantes isotípicos Los determinantes isotípicos establecen la diferencia entre las regiones constantes de las diversas clases y subclases de cadenas ligeras; representan diferentes genes de la región constante [ver tabla 1]. Por ejemplo, en un individuo existen cuatro isotipos de la cadena pesada de IgG: g1, g2, g3 y g4, que representan las subclases IgG1, IgG2, IgG3 e IgG4, respectivamente. Hay un sólo isotipo de la cadena ligera k y un isotipo de la cadena ligera l.
Cerca del 60 porciento de las inmunoglobulinas están formadas de cadenas pesadas y ligeras k y 40 porciento consisten en cadenas pesada y ligeras l. La inmunización con ciertos antígenos induce la formación de anticuerpos con la misma distribución de determinantes k y l, que la encontrada en la población general de inmunoglobulinas; sin embargo, algunos antígenos provocan la formación de inmunoglobulinas compuestas sobre todo por cadenas k o l; del mismo modo, algunos antígenos inducen más la producción de una clase en particular de inmunoglobulinas que otros. Se desconoce qué mecanismos determinan la clase de inmunoglobulinas que se producen o qué factores controlan el tipo de cadena ligera que se unirá a las cadenas pesadas.
Determinantes alotípicos Los determinantes alotípicos distinguen entre las inmunoglobulinas de un isotipo en particular, representan alelos diferentes de los genes de las inmunoglobulinas y, por lo tanto, se determinan genéticamente de acuerdo con las leyes mendelianas de manera muy similar a la que determina los grupos sanguíneos ABO. Las moléculas de cadena pesadas g de un individuo pueden tener varios marcadores diferentes, mientras que no se conocen marcadores alotípicos para las cadenas pesadas µ, d y e o para las cadenas ligeras l. Las cadenas pesadas g poseen más de 20 marcadores alotípicos distintos, que se determinan en forma colectiva Gm. Además, las cadenas ligeras k contienen un grupo de marcadores alotípicos llamados Inv. Se han reconocido tres de estos marcadores: Inv 1, Inv 2 e Inv 3.
Determinantes idiotípicos Un idiotipo se define como un determinante antigénico único localizado en la región variable de un anticuerpo. Un idiotipo es la expresión antigénica de la región variable de un anticuerpo. Los determinantes idiotípicos distinguen a una inmunoglobulina de otra del mismo alotipo. Si un animal es inmunizado con una proteína de mieloma (i.e., una inmunoglobulina homogénea producida por células plasmáticas neoplásicas) el anticuerpo resultante tendrá anticuerpos dirigidos contra muchas partes de la molécula de mieloma. Cuando se extraen los anticuerpos de este antisuero que reaccionan con otras proteínas de mieloma del mismo isotipo y alotipo, el antisuero resultante seguirá siendo capaz de reaccionar con las proteínas inmunogénicas del mieloma pero no con otras. Pueden realizarse experimentos similares utilizando anticuerpos monoclonales. Los antisueros reaccionan con determinantes localizados en la porción variable de las cadenas ligeras y pesadas (ver adelante). En situaciones en las que se encuentra que la proteína de mieloma es un anticuerpo dirigido contra un hapteno específico, la agregación de tal hapteno evitará que la proteína de mieloma reaccione con el suero anti-idiotípico [ver figura 7]. Esto indica que los determinantes idiotípicos están conectados de manera íntima con el sitio de unión del antígeno en la inmunoglobulina. Los anticuerpos inducidos por un antígeno único tendrán diferentes especificidades idiotípicas, lo que ilustra una vez más la gran heterogeneidad de las inmunoglobulinas.
|
| Figura 7 |
| Modelos murinos |
ESTRUCTURA Y SECUENCIA DE LAS INMUNOGLOBULINAS
Los estudios iniciales realizados con las proteínas de Bence Jones (dímeros de cadenas ligeras), con las proteínas de mieloma y con anticuerpos homogéneos purificados, produjeron un descubrimiento sorprendente: cuando se compararon anticuerpos de la misma clase, se encontró que una parte de cada cadena polipeptídica del anticuerpo era la misma, pero otra parte era muy variable. En la actualidad se sabe que la porción terminal carboxilo (C) de todas las cadenas ligeras k tiene una secuencia de aminoácidos idéntica, y por lo tanto a esta región se le denomina dominio constante o C, mientras que la porción terminal amino (N) tiene una secuencia de aminoácidos variable denominada secuencia V [ver figura 4]. Los primeros 110 aminácidos de la porción N-terminal de la cadena l ligera y de la cadena pesada también son variables, mientras que el 75 porciento restante de la cadena pesada es constante y consiste en un grupo de tres regiones homólogas. La semejanza entre las regiones constantes de las cadenas ligeras y pesadas indica que éstas pueden haber evolucionado a partir de un gen ancestral común.
Las regiones variables de las cadenas ligeras y pesadas pueden dividirse en varios subgrupos. Por ejemplo, con base en las diferencias en sus regiones variables, se distinguen cuatro subgrupos de cadenas ligeras k y seis subgrupos de cadenas ligeras l. Existen también tres subgrupos de regiones variables en las cadenas pesadas
Dentro de las regiones variables existen tres áreas que muestran gran variación [ver figura 8], y que se conocen como regiones hipervariables, correspondiendo al sitio de combinación del antígeno con el anticuerpo.
|
|
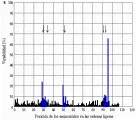

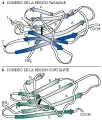
|
| Figura 9 |
| Estructura terciaria de las inmunoglobulinas |
La IgG puede dividirse en tres fragmentos por la papaína, una enzima preteolítica [ver figura 10]. Dos de estos fragmentos son similares y se denominan Fab; el tercer fragmento se denomina Fc. La porción Fc es responsable de la actividad biológica de las diversas inmunoglobulinas; entre otras funciones, la porción Fc controla la capacidad de las inmunoglobulinas para unirse a las células, fijar complemento y atravesar la placenta. Puede observarse que la papaína actúa sobre el extremo amino terminal de los puentes S-S que mantienen unidas a las cadenas pesadas entre sí.
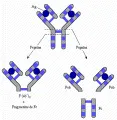
|
| Figura 10 |
| Sitios de separación de la inmunoglobulina |
Otra enzima proteolítica, la pepsina, divide a la molécula de IgG más allá de los puentes S-S que unen a las cadenas pesadas, dejando un fragmento de gran tamaño, F(ab')2, el cual tiene la capacidad de unir y precipitar antígenos por su valencia doble y por su capacidad para formar una malla. En este procedimiento, la digestión enzimática divide a la fracción Fc en muchos fragmentos pequeños. La porción que es susceptible a la digestión por ambas enzimas se denomina región de unión o de bisagra, las enzimas proteolíticas pueden tener acceso a esta región porque contiene muchos residuos de prolina que evitan que la cadena peptídica se pliegue y forme una hélice.
Los anticuerpos catalíticos son un tipo novedoso de inmunoglobulina.6,7 Estos anticuerpos monoclonales se originan contra haptenos que recuerdan el estado de transición de una reacción enzimática, como la hidrólisis del éster carboxílico. Los anticuerpos se comportan como enzimas cuando se mezclan con las sustancias adecuadas.
SINTESIS DE ANTICUERPOS (IN VITRO E IN VIVO)
Cuando el antígeno se introduce por primera vez al organismo, se presenta una respuesta primaria que se caracteriza por una fase latente, que dura varios días, durante la cual no se detectan anticuerpos; en seguida se produce un aumento en la cantidad en suero de anticuerpos IgM, que suelen alcanzar un nivel máximo después de siete días. Al cabo de seis a siete días, también se detectan anticuerpos IgG. El pico de IgM empieza a disminuir antes de que se alcance el título máximo de IgG después de 10 a 14 días; a continuación, los títulos de anticuerpos disminuyen y muy poco antígeno se detecta cuatro a cinco semanas después de una dosis única de antígeno. En un segundo encuentro con el antígeno se producirá una respuesta secundaria (también denominada respuesta anamnésica o de refuerzo). Tanto los títulos de IgM como los de IgG se elevan de manera exponencial, sin la fase latente que se observa en la respuesta primaria [ver figura 11]. Mientras que el pico de IgM puede ser de la misma magnitud o ligeramente mayor que el que se presenta durante la respuesta primaria, la respuesta de la IgG es mucho mayor y dura más tiempo. Esta variación en la respuesta es una demostración apropiada de la memoria inmunológica, y se debe a la proliferación de células B antígeno específicas y de células T cooperadoras durante la respuesta primaria. Las características de las respuestas primaria y secundaria explican la necesidad de inyecciones de refuerzo en los programas de inmunización.
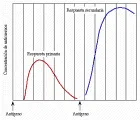
|
| Figura 11 |
| Respuesta al antígeno |
Maduración de la afinidad
Las propiedades de unión del anticuerpo cambian con el tiempo. A este proceso se le denomina maduración de la afinidad, e incluye el proceso de maduración somática y de selección. Después del estímulo primario, los anticuerpos tienen una afinidad cada vez mayor por el antígeno según va pasando el tiempo, y forman complejos antígeno-anticuerpo con una mayor estabilidad. Además, los anticuerpos se vuelven menos específicos y presentan más reacciones cruzadas con antígenos relacionados. En realidad, la disminución en la especificidad refleja el hecho de que las reacciones cruzadas, que previamente eran muy débiles para ser detectadas, se vuelven aparentes al tener el anticuerpo mayor afinidad por el antígeno.
Mecanismos de control
La IgG no se incrementa de manera indefinida, deben existir algunos mecanismos de control para detenerla. Si esto no ocurriera, las células proliferantes escaparían al control y saturarían al organismo con anticuerpos. De hecho, parecen existir varios mecanismos de control; primero, disminuye la cantidad de antígeno. Segundo, existen evidencias considerables de que la IgG específica contra antígeno suprime por sí misma la síntesis de IgG. Aunque puede suponerse en primer término que esta reacción se produce por el simple bloqueo del antígeno, esta explicación no es válida por completo debido a que la porción F(ab')2 de la IgG no suprime la síntesis de IgG de una manera tan eficaz como la molécula completa de IgG. Tercero, tal vez participe la regulación idiotípica [ver figura 7]. Por último, las células T supresoras también ejercen control sobre la producción de anticuerpos. De hecho, al encontrar títulos inexplicablemente elevados de IgE en seres humanos, debe considerarse la posibilidad de una falta de células T supresoras.
Competencia antigénica
Otro ejemplo del control de la produción de anticuerpos es el fenómeno de competencia antigénica. Si ciertos antígenos se administran de manera secuencial o combinados, un antígeno impedirá la producción de anticuerpos contra el segundo; por ejemplo, a menos que los toxoides tetánicos y diftérico sean administrados en la proporción correcta, un antígeno impedirá una respuesta inmunológica contra el otro. Así mismo, cuando se inyecta una inmunoglobulina (i.e., a un conejo) se presenta una elevación de anticuerpos mucho mayor contra la porción Fc que contra la porción Fab. Si se inyectan cantidades iguales de fragmentos Fab y Fc, entonces se producen anticuerpos contra los dos.
El análisis del fenómeno de la competencia antigénica sugiere que varios mecanismos se encuentran relacionados. Es probable que uno de éstos sea la producción, por las células T, de factores supresores no específicos. Puede suceder que el primer antígeno administrado o el antígeno dominante estimulen la producción de factores de las células T, los cuales eviten la respuesta contra el segundo antígeno. Este posible mecanismo de la competencia antigénica puede ser importante en la inmunosupresión ocasionada por virus. De manera adicional, en algunos casos existe un límite en el número de células B antígeno-específicas que pueden competir por los dos antígenos, lo cual es probablemente el mecanismo que limita a los anticuerpos contra la porción Fc de la molécula de la IgG cuando se inyecta la inmunoglobuliina completa.
Cambio en la producción
Como se mencionó antes, la producción de IgM va seguida por la producción de IgG. En la actualidad existe evidencia considerable de que las células productoras de IgM cambian hacia la producción de IgG [ver figura 12]; por ejemplo, la administración de anticuerpos dirigidos contra la cadena pesada de la IgM, la cadena µ, puede evitar la producción subsecuente tanto de IgM como de IgG, mientras que los anticuerpos contra la cadena pesada de la IgG, la cadena g, sólo evitan la síntesis de la IgG. Más aún, algunos pacientes tienen mielomas de las clases IgM e IgG y el análisis ha demostrado que estos mielomas comparten el mismo idiotipo. En otras palabras, las regiones variables de sus cadenas ligeras y pesadas son las mismas, pero las regiones constantes de sus cadenas pesadas difieren. Estas observaciones no sólo son de importancia para comprender la manera en que las células cambian de la producción de IgM a la producción de IgG, sino que también tienen implicaciones importantes para el control genético de la síntesis de inmunoglobulinas.
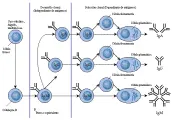
|
| Figura 12 |
| Desarrollo de células tronco |
INMUNOGLOBULINAS COMO RECEPTORES
Receptores de células B
Además de secretarse, las inmunoglobulinas pueden expresarse en la superficie de las células B, en donde actúan como receptores de antígenos.8 Estas inmunoglobulinas de superficie de membrana (IgSm) difieren de las inmunoglobulinas secretadas en que tienen un dominio transmembrana y son monoméricas. La primera IgSm que expresa una célula B es IgM, en una fase más avanzada del desarrollo se expresa en forma conjunta IgD [ver adelante, Origen genético de la diversidad de los anticuerpos]. La IsSm no viaja a la superficie celular por sí misma, sino que el proceso requiere la formación de un complejo consistente en la Ig y dos cadenas polipeptídicas denominadas Iga e Igb en el retículo endoplásmico [ver figura 13a]. El receptor de la célula B resultante se une a un antígeno. La célula B procesa el antígeno y expresa un fragmento peptídico en una molécula del CPH de clase II en la superficie celular para presentarlo a una célula T cooperadora. La estimulación de la célula T activa a la célula B, haciendo que se diferencíe a una célula de memoria o plasmática, que produce anticuerpos secretorios específicos para el antígeno encontrado. Después de la diferenciación terminal no se expresan más Iga o Igb. La célula plasmática madura cesa de expresar IgSm, a pesar de que mantenga el ARN mensajero (ARNm) para ésta.
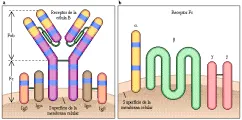
|
| Figura 13 |
| Receptores de la célula B y Fc |
Receptores para Fc
Se expresan receptores para Fc en múltiples células, incluyendo células cebadas, macrófagos, eosinófilos y células tumorales. Los receptores para Fc están compuestos de una familia de moléculas y su función es fijar la porción Fc de las inmunoglobulinas a la célula. El receptor de Fc para la IgE (RFce) sirve como modelo para todos los receptores de este tipo [ver figura 13b].9 Consiste en tres cadenas polipeptídicas, designadas como a, b, y g. La cadena RFce a es el sitio de unión para la porción Fc de la IgE. La RFceb es una molécula transmembrana que conecta a RFcea con la RFceg, la cadena responsable de reclutar las moléculas encargadas de las señales de transducción.
ORIGEN GENETICO DE LA DIVERSIDAD DE LOS ANTICUERPOS
El aspecto más intrigante del control genético de la síntesis de inmunoglobulinas es la diversidad del producto: las células plasmáticas pueden producir anticuerpos que reaccionen con más de 109 sitios antigénicos diferentes. ¿Cómo puede el ADN codificar contra un número tan grande de antígenos, muchos de los cuáles existen (en la escala de la evolución) desde hace muy poco tiempo?
La respuesta a esta pregunta ha empezado a surgir de los trabajos realizados utilizando técnicas de ADN recombinante.10-12 En estos estudios, el ADN de células embrionarias o germinales (espermatozoides) se comparó con el ADN, el ARN nuclear y el ARN mensajero (ARNm) de plasmocitomas. Los resultados indican, por ejemplo, que el ADN embrionario de la cadena ligera k codifica más de 300 regiones variables (V), cinco regiones de unión (J) y una región constante (C). Las regiones V y J se encuentran separadas del segmento C por una porción de ADN. Por lo tanto, el ADN embrionario tiene la capacidad para formar al menos 1,500 combinaciones diferentes de V y J; en otras palabras, por lo menos 1,500 cadenas ligeras k diferentes.
La aparición de estas líneas celulares individuales es el resultado de la recombinación somática en el ADN y de la unión del ARN [ver figura 14]. Al diferenciarse las células embrionarias en células plasmáticas, delecciones en el ADN hacen que uno de los genes V, escogidos al azar, sea adyacente a uno de los genes J. Esta unidad V-J y el resto de las regiones J están separadas del gen C por una porción corta de ADN. En el siguiente paso, el ADN es trancrito al ARN nuclear y la secuencia queda lista para un segundo rearreglo.
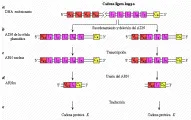
|
| Figura 14 |
| Linajes de células plasmáticas |
Este evento se inicia cuando una enzima rompe el ARN nuclear para producir ARN mensajero (ARNm). En un proceso denominado unión del RNA, el segmento que separa a la región o unidad V-J de la región C es removido junto con los segmentos J sobrantes. El segmento V-J-C restante se traduce en una de las 1,500 o más proteínas de las cadenas ligeras k. En realidad el número es mayor por el hecho de que la unión de cualquier porción V con J puede afectar alguna entre una variedad de secuencias de bases apareadas en el sitio de recombinación.
La variabilidad en la cadena pesada también contribuye de manera importante a la especificidad de un anticuerpo. Esta variabilidad es consecuencia de un proceso que incluye la recombinación somática y la unión de ARN, similares al proceso que crea la variabilidad en las cadenas ligeras. El ADN embrionario lleva las instrucciones para la formación de varios cientos de genes V de cadena pesada, seis regiones J, 10 a 20 regiones D (diversidad) y nueve genes C (estos genes C codifican las clases de cadenas pesadas: IgM, IgD, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2 e IgE). La deleción de ADN, su transcripción al ARN nuclear y la unión del ARN producen la secuencia V-C-J-J final en el ARNm, que es codificada por los ribosomas a una proteína de cadena pesada.
La operación aritmética se encuentra ahora completa: puede calcularse que el mecanismo ADN-ARN de cadena pesada produce más de 18,000 variedades posibles (la especificidad del anticuerpo no varía con la clase del mismo, por lo que los nueve genes C no entran en el cálculo). La combinación de las más de 1,500 variedades de cadenas ligeras con las 18,000 variedades de cadenas pesadas podría producir más de 27 millones de anticuerpos diferentes con sitios de combinación para antígenos diferentes.
De esta manera, parecería que la diversidad de los anticuerpos en las cadenas ligeras k y en las cadenas pesadas puede ser producida por la recombinación somática de los genes de las líneas germinales. Sin embargo, esta no es la única base para la generación de diversas moléculas de anticuerpos. Los estudios realizados en la cadena ligera l indican un mecanismo adicional. Estas cadenas son muy similares entre sí y sólo muestran variación ocasional en su secuencia de nucleótidos. En vista de que sólo se ha encontrado un segmento Cl1 por genoma haploide, sólo las mutaciones en punto pueden explicar este hallazgo. Se han encontrado variantes somáticas semejantes en las cadenas ligeras k y en las cadenas pesadas. La frecuencia de mutación somática es relativamente elevada (un par de bases por cada 1,000 divisiones celulares).
Las secuencias cortas y muy conservadas de ADN que limitan los segmentos V, D y J son utilizadas por la célula como señales para el rearreglo de los genes de las inmunoglobulinas [ver figura 15].10,12 Estas secuencias de ADN consisten en siete pares de bases (heptámero) y de una secuencia de nueve pares de bases (nonámero) separadas por 12 ó 23 pares de nucleótidos al azar, que constituyen los nucleótidos de dispersión. Una vuelta de la hélice del ADN equivale aproximadamente a doce pares de bases; por lo tanto, las secuencias que contienen nucleótidos de dispersión de doce pares de bases se denominan señales de una vuelta. En forma análoga, las secuencias que contienen nucleótidos de dispersión de 23 pares de bases, equivalentes a dos vueltas de la hélice de ADN, se denominan señales de dos vueltas. La recombinación de los segmentos de los genes de las inmunoglobulinas parece depender del apareamiento de estas señales de recombinación complementaria; sin embargo, al parecer el apareamiento sólo ocurre entre las señales de una de dos vueltas (la regla de unión de una o dos vueltas). La distribución de los dos tipos de secuencias de recombinación garantiza que sólo puedan ocurrir ciertas recombinaciones de genes.
|
|
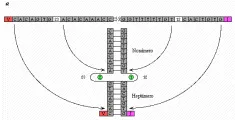
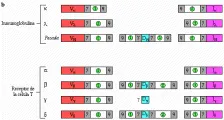
Los genes que codifican a las cadenas pesadas de IgM e IgD (los genes µ y d) desempeñan una función muy importante en la respuesta primaria, que ocurre como resultado de la primera exposición al antígeno. Las células B iniciales expresan sólo IgM sobre su superficie; más tarde en la ontogenia, se expresa la IgM en la membrana celular junto con el producto final del gen de IgD. La IgM aumenta durante la ontogenia, alcanzando concentraciones en las células plasmáticas completamente diferenciadas alrededor de 100 veces más altas que las concentraciones en las células pre-B. Cuando las células B se diferencían, la IgD unida a la membrana disminuye hasta una concentración aproximada del 10 porciento de la IgM unida a la membrana. Junto con estos cambios cuantitativos ocurre una mayor modulación del desarrollo de las formas secretadas y unidas a la membrana de cada cadena pesada. Las formas de membrana de IgM e IgD predominan en las etapas iniciales en vista de que constituyen receptores para antígenos específicos. Sin embargo, ocurre un cambio al final de la diferenciación que ocasiona la producción casi exclusiva de la forma secretada de IgM. Aunque la IgD es un isotipo abundante en la superficie celular, es secretada sólo en pequeñas cantidades.15,16
En el cambio de producción de IgM a IgG que constituye la respuesta de refuerzo, los genes de la región constante son separados del ADN de la célula plasmática antes de que ocurra la transcripción al ARN.17,18 Por ejemplo, si la célula cambia hacia la producción de IgG3, los genes para µ y d son suprimidos [ver figura 16]. Después de la transcripción, la unión del ARN produce un mensajero con la secuencia V-D-J-Cg3, la cual se traduce a la proteína.
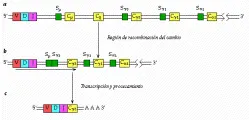
|
| Figura 16 |
| Respuesta inmune secundaria |
HIBRIDOMAS DE LINFOCITOS
El desarrollo del hibridoma de linfocitos, productos de fusión celular, tiene implicaciones revolucionarias para la inmunología y la medicina clínica.19 Los hibridomas de linfocitos constituyen el medio por el cual pueden producirse títulos muy altos de anticuerpos puros, muy específicos, para fines experimentales. Con este recurso los inmunólogos han sido capaces de incrementar y profundizar el conocimiento de las funciones del sistema inmune. Por ejemplo, los anticuerpos monoclonales han ayudado a definir los productos genéticos del CPH y muchos marcadores de superficie de las células hematopoyéticas, y han sido importantes para estudiar las bases de la mutación somática en los genes de los anticuerpos. En el campo clínico, la producción de estos anticuerpos ha permitido realizar pruebas serológicas de mucha mejor calidad. Se han usado anticuerpos monoclonales para eliminar células T y células tumorales antes del trasplante de médula ósea, y también en receptores de trasplante para evitar el rechazo de riñón o hígado al disminuir el número de células T citotóxicas.
El hibridoma de linfocitos, tal y como fue desarrollado por Köhler y Milstein, es el producto de la fusión de una célula de mieloma de ratón y un linfocito obtenido del bazo de un ratón inmunizado con un antígeno específico. La fusión puede lograrse empleando una variedad de virus o polietilenglicol. El resultado es una célula única con dos núcleos. Después, la membrana nuclear se rompe y los cromosomas se dividen y se distribuyen al azar entre las dos células hijas, los hibridomas de linfocitos.
Estas manipulaciones se llevan a cabo en un medio selectivo. Las células de mieloma, las cuales son incapaces de crecer en este medio, debido a que carecen de las enzimas necesarias, mueren. Aunque las células esplénicas poseen esta enzima, también mueren, debido a que no son capaces de replicarse de manera indefinida; sin embargo, las células híbridas tienen tanto la enzima del linfocito como la capacidad de la célula de mieloma para sobrevivir de manera indefinida, de manera que viven y al hacerlo inmortalizan la capacidad de la célula esplénica para elaborar anticuerpos específicos.
Los híbridos pueden crecer y ser clonados. El medio de cultivo de las clonas se prueba en busca de anticuerpos contra el antígeno deseado, de manera que las clonas positivas pueden ser seleccionadas y cultivadas en masa, in vitro o in vivo. De cualquier manera, debido a que se originan de una clona única, las células productoras de anticuerpos elaboran un solo tipo de anticuerpo en grandes cantidades. En comparación, las técnicas clásicas producen sueros que contienen un amplio espectro de anticuerpos diferentes al deseado, aun cuando la titulación sea muy elevada.
Un ratón que tenga hibridomas de linfocitos producirá anticuerpos específicos (por ejemplo, contra un antígeno de superficie linfocitario), en una cantidad mayor que todos los anticuerpos contra este antígeno sintetizados por muchos investigadores en todo el mundo. Tal anticuerpo puede tener títulos por millones, un fenómeno sin precedentes; así mismo, debido a que el anticuerpo se dirige contra un antígeno único, puede ser empleado para aislar ese antígeno.
ANTICUERPOS MONOCLONALES HUMANOS
En la actualidad existen varios métodos para elaborar anticuerpos monoclonales humanos. El primero consiste en tomar el ADN complementario que codifica un anticuerpo monoclonal murino y remplazar en forma sistemática las secuencias murinas con secuencias humanas. El ADN que codifica para las regiones constantes de las cadenas pesadas y ligeras humanas remplaza las secuencias de ADN murinas de las regiones variables con humanas por medio de mutagénesis, dejando las regiones hipervariables intactas. Una desventaja de este método es que el marco de lectura de las regiones variables puede afectar la afinidad. Por lo tanto, las inmunoglobulinas humanizadas pueden requerir mutaciones adicionales para aumentar su afinidad.
Otro método de humanizar anticuerpos monoclonales murinos consiste en tomar un segmento grande de ADN humano que incluya varias de las regiones variables humanas y todas las regiones constantes humanas e introducir este segmento en el genoma murino. Este ratón transgénico se cruza con un ratón en el que exista ruptura dirigida de la región J de los genes de las cadenas pesadas de Ig y ligeras k.20 Puede entonces inyectarse cualquier antígeno al ratón transgénico, y producirá anticuerpos monoclonales humanizados. Debido a que el ratón transgénico contiene un número limitado de regiones variables humanas, la potencia de estos anticuerpos depende de la mutación somática natural y de la maduración de la afinidad que ocurra en el ratón.
Un tercer método de generar anticuerpos monoclonales humanos consiste en construir bibliotecas (inventarios) de regiones variables humanas en bacterias o en bacteriófagos. El anticuerpo puede expresarse en la superficie de la bacteria o el bacteriófago y seleccionarse por afinidad al antígeno de interés.
Los anticuerpos monoclonales son de gran utilidad para investigar los mecanismos de la respuesta inmune. Por ejemplo, se usan para aislar antígenos de la superficie celular y definir subgrupos de linfocitos o de otras células. Son un instrumento ideal para producir grandes cantidades de anticuerpos homogéneos para estudiar la estructura fina del sitio de combinación del anticuerpo con un antígeno específico. También se han usado los hibridomas de linfocitos para determinar la base genética de la generación de la diversidad en la formación de anticuerpos. Administrados in vivo, los anticuerpos monoclonales permiten investigar el papel de varios subtipos de linfocitos que regulan el sistema inmune, al eliminarlos o activarlos. También se emplean para neutralizar diversas citocinas y para definir su función en la inmunorregulación e inflamación.
Los anticuerpos monoclonales pueden ser muy útiles en otros campos además de la inmunología. Por ejemplo, los anticuerpos contra los receptores hormonales pueden ser capaces de unirse al receptor y modificar una respuesta endócrina.
Receptores de células T
A diferencia de los receptores de inmunoglobulinas de las células B, que pueden reconocer antígenos libres, los receptores de células T sólo reconocen antígenos combinados con antígenos autólogos del complejo principal de histocompatibilidad, por lo general sobre la superficie de los macrófagos. Las células T cooperadoras o CD4+ (así como las pocas células T citotóxicas CD4+), requieren moléculas clase II del CPH, y las células T citotóxicas CD8+ requieren moléculas clase I del CPH. Este fenómeno se conoce como restricción del CPH. La capacidad de las células T para reconocer estas moléculas del CPH propias se establece en el timo antes de que los linfocitos se expongan al antígeno .
Las moléculas que forman el receptor de la célula T y los genes que codifican estas moléculas se han aislado y se están definiendo mediante el uso de anticuerpos monoclonales que reconocen las clonas de células específicas (clonotípicas), hibridomas y linfomas de células T21-23 y con el empleo de tecnología de ADN recombinante.24-27 Existen datos convincentes de que el receptor de las células T está compuesto de seis polipéptidos distintos conocidos como complejo CD3-receptor de células T (RCT-CD3)28 [ver figura 17]. Del 85 al 95 porciento de los linfocitos de la sangre periférica tienen receptores RCT-ab y sólo el cinco al 15 porciento tienen RCT-gd. Parece que los receptores gd son expresados por un subgrupo de linfocitos derivados del timo diferentes a las células que expresan RCT-ab. La porción del complejo RCT-ab que reconoce al antígeno está compuesta de dos cadenas polipeptídicas glucosiladas, denominadas RCT-a (45-55 kd) y RCT-b (37-45 kd), que están unidas por puentes disulfuro, formando un heterodímero de 90 kd. Las cadenas polipeptídicas correspondientes en el complejo RCT-gd consisten en RCT-g (45 a 55 kd)29 y RCT-d (35 a 45 kd). El mapeo de péptidos y la identificación de la secuencia de los aminoácidos de los heterodímeros en clonas de linfocitos con diferentes especificidades antigénicas, muestran que cada una de las cadenas de RCT-a, RCT-b, RCT-g y RCT-d contienen porciones variables (V) y constantes (C) análogas a las observadas en las inmunoglobulinas. No resulta sorprendente la presencia de regiones variables en moléculas que deben reconocer diversos antígenos. Las células RCT-ab pueden funcionar como células cooperadoras o como células asesinas, pero la única función relacionada con las células RCT-gd es la citotoxicidad. Se ha demostrado que algunas células RCT-gd producen lesiones cutáneas en la lepra.30
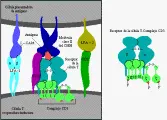
|
| Figura 17 |
| Receptor célula T/complejo CD3 |
Los heterodímeros de RCT-ab y RCT-gd están muy relacionados con las proteínas CD3: CD3-g (25 a 28 kd), CD3-d (20 kd), Cd3-e (20 kd) y CD3-s (16 kd) [ver figura 16]. Se ha utilizado un grupo de anticuerpos monoclonales, incluyendo OKT3, anti-Leu-4 y UCHT1, para identificar las proteínas CD3 (con mayor frecuencia (CD3-e) en todas las células periféricas de sangre periférica y en el 30 al 60 porciento de los timocitos. Todos los demás timocitos, excepto un pequeño porcentaje, expresan una baja concentración de CD3 y RCT en su superficie.
Al llegar al timo para convertirse en una célula T, el pretimocito expresa genes CD3 de inmediato, pero solo en el interior de la célula. Este timocito CD3-, RCT- en la superficie (CD3+ en el citoplasma) se desarrolla en un compartimiento del timo que no tiene moléculas de superficie CD4 y CD8, por lo que se denomina timocito CD4-, CD8- o doble negativo (DN). En un subcompartimiento de los timocitos DN ocurre rearreglo y expresión del locus RCT-b (determinado por los genes rag1 y rag2, además de otros genes desconocidos). Por el contrario, el rearreglo y expresión del RCT-a ocurre en una etapa más tardía del desarrollo, en un compartimiento en el que los timocitos expresan moléculas de superficie tanto CD4 como CD8 para volverse timocitos CD4+, CD8+ o doble positivas (DP). El complejo RCT-CD3 se expresa completo en la superficie celular solo hasta esta fase del desarrollo de los timocitos DP. Una vez expresado, el complejo RCT-CD3 es responsable del proceso de selección positiva o negativa. Para la selección positiva el complejo RCT-CD3 tiene que reaccionar con un fragmento peptídico unido a una molécula de clase I del CPH para producir una célula T CD8+ o con un fragmento peptídico unido a una molécula de clase II del CPH para producir una célula T CD4+, ambas conocidas como células sencillas positivas. Esta vía de desarrollo ha sido confirmada por mutaciones del RCT producidas en ratones en los que se han eliminado genes específicos [ver figura 18]. Los ratones en los que se eliminaron los genes rag1, rag2 u otros que gobiernan la expresión del locus RCT-b presentaron un bloqueo en el desarrollo de los timocitos a nivel del timocito DN.31 Por el contrario, la eliminación del gen del RCT-a causa bloqueo del timocito DP. La alteración de los genes CD3 provoca bloqueo parcial del desarrollo de los timocitos por la redundancia de los componentes del CD3.
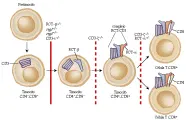
|
| Figura 18 |
| Vía de desarrollo de una célula T |
La expresión de las proteínas CD3 y RCT sobre la superficie celular es dependiente en forma mutua: ninguno de los componentes se observa sobre la superficie de las células T sin el otro. Por ejemplo, las células mutantes que no producen transcritos de ARNm de la cadena RCT-b carecen por completo de las proteínas de superficie CD3 y RCT; la transferencia de una copia normal del gen de la RCT-b en estas células restablece la transcripción y produce un nivel normal de las proteínas CD3 y RCT sobre la superficie celular. Además, la incubación de células T con anticuerpos dirigidos contra el RCT o el CD3 (o la incubación de células T antígeno-específicas con péptidos antigénicos específicos) causa comodulación, esto es, la desaparición del RCT y el CD3 de la superficie celular y su aparición en cuerpos multivesicales que más tarde se fusionan con lisosomas. La comodulación se acompaña de la pérdida de funciones antígenoespecíficas. El RCT y el CD3 se vuelven a expresar alrededor de 49 horas después de extraer los anticuerpos moduladores.
Los datos estructurales y funcionales indican que las actividades del receptor de células T se distribuyen entre las subunidades del complejo RCT-CD3: los polipéptidos del RCT (a, b, g y d) unen al antígeno y a los productos del gen del CPH, y las proteínas CD3 traducen la señal de unión al citoplasma de la célula T, lo que produce la activación de las funciones de la célula T.
RECEPTORES DE CELULAS T Y GENES CD3
La organización de los genes que codifican las cadenas a, b, g, y d del receptor de las células T humanas [ver figura 19] es análoga a la de los genes de las cadenas pesadas de las inmunoglobulinas; existen segmentos variables, de diversidad y de unión, que están limitados por secuencias de reconocimiento que modulan la recombinación antígeno específica [ver antes, Anticuerpos].32,33 De este modo, la diversidad de los receptores de las células T se genera en gran parte por los mismos mecanismos utilizados por las células B en la producción de las inmunoglobulinas.34 De hecho, las células T y B utilizan la misma enzima de recombinación, o recombinasa.34
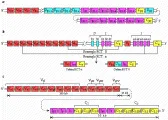
|
| Figura 19 |
| Locus del RCT-beta |
Las secuencias genómicas que codifican la cadena RCT-b contienen dos genes de la región constante muy similares, Cb1 y Cb2, cada uno de los cuales se asocia con un grupo de seis o siete secuencias J y una secuencia D única [ver figura 19a]. Existen al menos 70 genes Vb que se relacionan con dos genes Cb. El rearreglo de los segmentos del gen de la cadena b permite la producción de aproximadamente 3,600 cadenas b diferentes.
Los genes del receptor a de las células T están distribuidas en forma diferente [ver figura 19b]. Un sólo gen Ca está precedido por una cadena muy grande de ADN compuesta de por lo menos 50 secuencias J distintas. No se ha demostrado de manera directa la presencia de un segmento del gen Da. Algunas secuencias Va se organizan como familias de genes relacionados. El rearreglo de los segmentos del gen de la cadena a puede representar cerca de 2,500 polipétidos diferentes. Por lo tanto, se pueden formar 9 X 106 RCTab. Aunque no se han detectado mutaciones somáticas en los receptores de células T, la recombinación imprecisa contribuye a la variabilidad de las cadenas RCT-a y RCT-b. Por consiguiente, la cantidad de RCT-ab puede ser considerablemente mayor de 9 X 106.
Los genes de cadenas RCR-b y RCT-g se han localizado en el cromosoma 7 y los genes de las cadenas RCT-a y RCT-d en el cromosoma 14, mediante el uso de híbridos de células somáticas de roedores y humanos y por técnicas de hibridación in situ [ver tabla 3]. Los genes para RCT-b y RCT-g se han localizado en brazos opuestos del cromosoma 7. Estos estudios de mapeo cromosómico indican que los genes del receptor de células T no están ligados al complejo principal de histocompatibilidad. Aunque existe cierta superposición en la localización cromosómica de los genes del receptor de células T y de las inmunoglobulinas, no hay consistencia entre las especies: los genes de la RCT-a y de las cadenas pesadas de las inmunoglobulinas se mapean en el cromosoma 14 en humanos pero se encuentran en cromosomas separados en el ratón, y los genes de las cadenas k de las inmunoglobulinas y de la RCT-b se localizan en el cromosoma 6 en el ratón pero no están relacionados en el genoma humano [ver tabla 3].
|
||||
RCT: Receptor de células T. CD3: Antes denominado T3.
|
Los genes CD3-g, CD3-d CD3-e y CD3-s se transcriben en todas las células T, pero no se rearreglan. El CD3-g, CD3-d y CD3-e son codificados en la porción distal del brazo largo del cromosoma 11 humano (11q23-11qter); el CD3-s se localiza en el cromosoma 1 [ver tabla 3]. Los genes CD3-g, CD3-d y CD3-e se agrupan en una zona del cromosoma 11 con una extensión de cerca de 50,000 pares de bases. Su expresion específica en las células T es regulada por lo menos por dos factores potenciadores localizados justo hacia adelante de los genes CD3-d y CD3-e.35,36 Se considera que los genes CD3 desempeñan una función crucial en el inicio de la diferenciación intratímica. Estos genes de las cadenas de CD3 no están relacionados con los genes del CPH, del RCT o de las inmunoglobulinas, ni en humanos ni en ratones.
Respuesta inmune humana en modelos animales
Los ratones transgénicos son muy útiles en el estudio de la expresión del gen del receptor de las células T debido a un fenómeno conocido como exclusión alélica, que también ocurre en los genes de las inmunoglobulinas. La exclusión alélica se refiere al hecho de que una vez que ocurre el rearreglo del gen del RCT en un cromosoma, se inhibe el rearreglo subsecuente en el otro cromosoma. Por consiguiente, cuando se introduce un gen RCT-a o RCT-b rearreglado exógenamente en las células germinales de un ratón, se inhibe de manera notable el rearreglo endógeno de los genes RCT-a o RCT-b. Los estudios realizados en ratones transgénicos han demostrado que los mecanismos para la selección de autoantígenos (específicamente las moléculas de las clases I y II del CPH) desempeñan una función importante en la diferenciación final de las células T. Por ejemplo, en una serie de experimentos se aislaron los genes RCT-a y RCT-b de la clase I del CPH de una clona de células T citotóxicas CD4-, CD8+ específica para el antígeno H-Y masculino.37,38 Estos genes RCT-a y RCT-b antígeno-específicos rearreglados, fueron inyectados enseguida en los óvulos fertilizados murinos; la progenie que incorporó en su genoma el ADN inyectado (i.e., ratones transgénicos) fue utilizada para realizar más estudios.
El análisis de los timocitos y de las células T de ratones transgénicos demostró que la interacción específica entre el receptor de las células T sobre la superficie celular de timocitos inmaduros y los antígenos tímicos de la clase I del CPH determinaba las etapas finales de la diferenciación. Al parecer, las células T autorreactivas son eliminadas por selección negativa, ya que en los ratones varones, las células T autoespecíficas (i.e., específicas para el antígeno H-Y y restringidas por moléculas específicas de la clase I del CPH) son suprimidas en el timo. Varios estudios realizados en ratas transgénicas (sin el antígeno H-Y) aportaron datos acerca del reconocimiento intratímico de los antígenos de la clase I del CPH y demostraron que la selección positiva también afectaba la diferenciación de las células T inmaduras. En primer lugar, la frecuencia relativa de los timocitos CD4-, CD8+ fue mayor en los ratones transgénicos que en la especie del ratón original o en otros ratones que portaban alelos clase I del CPH diferentes. En segundo lugar, sólo las células T CD4-, CD8+ expresan altas concentraciones de cadenas polipeptídicas transgénicas RCT-a y RCT-b sobre su superficie celular. En tercer lugar, en los experimentos realizados para reconstruir la población celular de timos con células tronco de ratones transgénicos RCT-ab, ratones transgénicos RCT-b, y ratones normales, sólo las células madre transgénicas RCT-ab se asociaron con una proporción elevada de células CD4-, CD8+. Por lo tanto, parece que la interacción específica entre el receptor de las células T sobre los timocitos inmaduros CD4-, CD8+ y el antígeno de la clase I del CPH, provoca la diferenciación de estos timocitos en células T maduras CD4, CD8+. Otros experimentos indican que el reconocimiento de los antígenos de la clase II del CPH controla la diferenciación de células CD4-, CD8+ en células T maduras CD4-, CD8+. Por consiguiente, los modelos experimentales con ratones transgénicos apoyan la hipótesis de que los mecanismos de selección positiva y negativa intervienen en la diferenciación de timocitos humanos inmaduros en células T CD4+ y CD8+ con capacidad funcional.
El ratón "humano" con inmunodeficiencia combinada severa (ratón con IDCS-hu) es un modelo murino experimental utilizando para el análisis de la diferenciación y de la función hematolinfoide humana, pero también constituye una forma más directa para estudiar el sistema inmune humano que el modelo con ratones transgénicos.39-41 El ratón con IDCS tiene una deficiencia muy importante de linfocitos; las células de la población B o T no pueden detectarse por medio de estudios serológicos o funcionales. Al injertar células hematopoyéticas hepáticas, tejido del timo y de los ganglios linfáticos de fetos humanos al ratón con IDCS, se observó diferenciación de células humanas T y B maduras. Se ha detectado un aumento temporal de células T CD4+ y CD8+ humanas, además de IgG humana, en la circulación periférica de estos ratones con IDCS-hu. Las células T CD4+ humanas del ratón con IDCS-hu pueden ser infectadas por el virus del SIDA.42 El modelo de la IDCS-hu tendrá múltiples aplicaciones en el estudio de la respuesta inmune y de las enfermedades inmunológicas humanas.
Bibliografía
-
Guillet J-G, Lai M-Z, Briner TJ, et al: Immunological self,
nonself discrimination. Science 235:865, 1987
-
Townsend ARM, Bastin J, Gould K, et al: Cytotoxic T lymphocytes
recognize influenza haemagglutinin that lacks a signal sequence. Nature
324:575, 1986
-
Beckman EM, Porcelli SA, Morita CT, et al: Recognition
of a lipid antigen by CD1-restricted ab+
T cells. Nature 372:691, 1994 [PMID
7527500]
-
Furlong ST, David JR: Adjuvants. Therapeutic Immunology,
1st ed. Austen KF, Burakoff SJ, Rosen FS, et al, Eds. Blackwell Scientific
Publications, Cambridge, Massachusetts (in press)
-
Afonso LC, Scharton TM, Vieira LQ, et al: The adjuvant effect
of interleukin-12 in a vaccine against Leishmania major. Science
263:235, 1994 [PMID
7904381]
-
Tramontano A, Janda KD, Lerner RA: Catalytic antibodies.
Science 234:1566, 1986 [PMID
3787261]
-
Pollack SJ, Jacobs JW, Schultz PG: Selective chemical
analysis by an antibody. Science 234:1570, 1986 [PMID
3787262]
-
Reth M: Antigen receptors on B lymphocytes. Annu Rev Immunol
10:97, 1992
-
Hulett MD, Hogarth PM: Molecular basis of Fc receptor function.
Adv Immunol 57:1, 1994
-
Sakano H, Huppi K, Heinrich G, et al: Sequence at the
somatic recombination sites of immunoglobulin light-chain genes. Nature
280:288, 1979 [PMID
111144]
-
Early P, Huang H, Davis M, et al: An immunoglobulin heavy
chain variable region is generated from three segments of DNA: VH,
D, and JH. Cell 19:981, 1980 [PMID
6769593]
-
Max EE, Seidman JG, Leder P: Sequences of five potential
recombination sites encoded close to an immunoglobulin k
constant region gene. Proc Natl Acad Sci USA 76:3450, 1979
-
Oettinger MA, Schatz DG, Gorka C, et al: RAG-1 and RAG-2,
adjacent genes that synergistically activate V(D)J recombination. Science
248:1517, 1990 [PMID
2360047]
-
Shinkai Y, Rathbun G, Lam KF, et al: RAG-2-deficient mice
lack mature lymphocytes owing to inability to initiate V(D)J rearrangement.
Cell 68:855, 1992 [PMID
1547487]
-
Word CJ, White MB, Kuziel WA, et al: The human immunoglobulin
Cm -Cd locus: complete
nucleotide sequence and structural analysis. Intl Immunol 1:296, 1989
-
Kuziel WA, Word CJ, Yuan D, et al: The human immunoglobulin
Cm -Cd locus: regulation
of m and d RNA expression
during B cell development. Intl Immunol 1:310, 1989
-
Hofker MH, Walter MA, Cox DW: Complete physical map of the
human immunoglobulin heavy chain constant region gene complex. Proc Natl
Acad Sci USA 86:5567, 1989 [PMID
2501792]
-
Berman JE, Mellis SJ, Pollock R, et al: Content and organization
of the human Ig VH locus: definition of three new VH
families and linkage to the Ig CH locus. EMBO J 7:727, 1988
[PMID
3396540]
-
Kohler G, Milstein C: Continuous cultures of fused cells
secreting antibody of predefined specificity. Nature 256:495, 1975 [PMID
1172191]
-
Lonberg N, Taylor LD, Harding FA, et al: Antigen-specific
human antibodies from mice comprising four distinct genetic modifications.
Nature 368:856, 1994 [PMID
8159246]
-
T cell receptor genes. Owen M, Simpson E, Eds. Oxford University
Press, New York (in press)
-
Kappler J, Kubo R, Haskins K, et al: The mouse T cell receptor:
comparison of MHC-restricted receptors on two T cell hybridomas. Cell 34:727,
1983 [PMID
6605198]
-
Alt F: Immunoglobulin genes. Molecular Immunology (Frontiers
in Molecular Biology series), 2nd ed. Hames BD, Glover DM, Eds. IRL Press,
Washington, DC, 1995
-
Yanagi Y, Yoshikai Y, Leggett K, et al: A human T cell-specific
cDNA clone encodes a protein having extensive homology to immunoglobulin
chains. Nature 308:145, 1984 [PMID
6336315]
-
Hedrick SM, Cohen DI, Nielsen EA, et al: Isolation of cDNA
clones encoding T cell-specific membrane-associated proteins. Nature 308:149,
1984 [PMID
6199676]
-
Brenner MB, McLean J, Scheft H, et al: Two forms of the T-cell
receptor g protein found on peripheral blood
cytotoxic T lymphocytes. Nature 325: 689, 1987 [PMID
3102967]
-
Modlin RL, Pirmez C, Hofman FM, et al: Lymphocytes bearing
antigen-specific gamma delta T-cell receptors accumulate in human infectious
disease lesions. Nature 339: 544, 1989 [PMID
2471938]
-
Mombaerts P, Iacomini J, Johnson RS, et al: RAG-1-deficient
mice have no mature B and T lymphocytes. Cell 68:869, 1992 [PMID
1547488]
-
Lefranc MP, Chuchana P, Dariavach P, et al: Molecular mapping
of the human T cell receptor gamma (TRG) genes and linkage of the variable
and constant regions. Eur J Immunol 19:989, 1989
-
Lai E, Concannon P, Hood L: Conserved organization of the
human and murine T-cell receptor b-gene families.
Nature 331:543, 1988 [PMID
2829029]
-
Yancopoulos GD, Blackwell TK, Suh H, et al: Introduced T
cell receptor variable region gene segments recombine in pre-B cells: evidence
that B and T cells use a common recombinase. Cell 44:251, 1986 [PMID
3484682]
-
Clevers H, Lonberg N, Dunlap S, et al: An enhancer located
in a CpG-island 3¢ of the TCR/CD3-e
gene confers T lymphocyte-specificity to its promoter. EMBO J 8:2527, 1989
[PMID
2583122]
-
Georgopoulos K, Van den Elsen P, Bier E, et al: A T cell-specific
enhancer is located in a DNase I-hypersensitive area at the 3¢
end of the CD3-d. EMBO J 7:2401, 1988 [PMID
2847918]
-
Bluthmann H, Kisielow P, Uematsu Y, et al: T-cell-specific
deletion of T-cell receptor transgenes allows functional rearrangement
of endogenous a- and b-genes.
Nature 334:156, 1988 [PMID
3260351]
-
Teh HS, Kisielow P, Scott B, et al: Thymic major histocompatibility
complex antigens and the ab T-cell receptor
determine the CD4/CD8 phenotype of T cells. Nature 335:229, 1988 [PMID
2970593]
-
McCune JM, Namikawa R, Kaneshima H, et al: The SCID-hu mouse:
murine model for the analysis of human hematolymphoid differentiation and
function. Science 241:1632, 1988 [PMID
2971269]
-
Mosier DE, Gulizia RJ, Baird SM, et al: Transfer of a functional
human immune system to mice with severe combined immunodeficiency. Nature
335:256, 1988 [PMID
2970594]
-
Kamel-Reid S, Dick JE: Engraftment of immune-deficient mice
with human hematopoietic stem cells. Science 242:1706, 1988 [PMID
2904703]
-
Namikawa R, Kaneshima H, Lieberman M, et al: Infection of
the SCID-hu mouse by HIV-1. Science 242:1684, 1988 [PMID
3201256]