Contenido del artículo
II ANTIGENOS, ANTICUERPOS Y RECEPTORES DE CELULAS T
DR. COX TERHORST, PH.D.
DR. JOHN DAVID
Un antígeno es cualquier sustancia capaz de generar una respuesta inmune, esto es, de reaccionar con células T y células B para inducir la formación de anticuerpos y linfocitos sensibilizados, que reaccionan con esos anticuerpos y células una vez que son formados. Los primeros antígenos estudiados fueron diversas proteínas extrañas y de microrganismos, y sigue siendo válido que las proteínas son antigénicas en forma casi universal. Las bases de la inmunogenicidad de las proteínas se desconocen, pero es probable que se relacionen con su configuración única y estable. En la actualidad se sabe que en condiciones apropiadas un amplio rango de moléculas pueden inducir una respuesta inmune y que en situaciones anormales el organismo incluso desarrollará una respuesta inmune contra sus propios antígenos.
Los polisacáridos pueden inducir la formación de anticuerpos cuando se acoplan con proteínas, y algunos polisacáridos purificados son antígenos eficaces por sí solos. Un ejemplo lo constituye el polisacárido purificado del neumococo, que puede utilizarse como vacuna contra la cepa de neumococo en particular de la que se obtuvo el polisacárido. Se ha demostrado que los anticuerpos humanos contra los polisacáridos son sobre todo de la clase IgG, aunque también se detectan cantidades menores de IgM e IgA [ver adelante, Clasificación de las inmunoglobulinas]. Aunque la mayor parte de los antígenos son macromoléculas, algunas moléculas pequeñas también son antigénicas. En los pacientes con lupus eritematoso generalizado, enfermedad de Crohn o esclerosis lateral amiotrófica ocurren anticuerpos contra ADN o ARN.
Los antígenos no sólo son reconocidos por anticuerpos sino también a través de receptores específicos de células B (RCB) [ver adelante, receptores de células B] y receptores de células T (RCT) [ver adelante, receptores de células T], que se localizan en las superficies extracelulares de las células B y T, respectivamente. En general, los RCT y las inmunoglobulinas reconocen diferentes determinantes antigénicos.
Existen dos tipos diferentes de receptores de células T: RCT-ad y RCT-gd. Los diversos tipos de células T tienen mecanismos diferentes para reconocer los antígenos. Por ejemplo, los RCT-ab reconocen antígenos que han sido procesados por células presentadoras de antígenos (CPA) para convertirse en fragmentos peptídicos unidos a moléculas del complejo principal de histocompatibilidad (CPH) de clase I o clase II en la superficie de las CPA. En contraste, los RCT-gd parecen no requerir la presentación de antígeno por el CPH. Las células T cooperadoras solo reconocen fragmentos peptídicos unidos a moléculas del CPH de clase II, mientras que las células T citotóxicas reconocen antígenos procesados virales presentados tanto en moléculas de clase I como II del CPH sobre la superficie de las células humanas infectadas por los virus. Se ha demostrado que los lípidos puros de micobacterias pueden presentarse como antígenos al RCT-ab por las moléculas CD1 en lugar de las moléculas clase I o II del CPH.
Los adyuvantes son sustancias capaces de aumentar la inmunogenicidad de los antígenos y son de gran importancia para la producción de las vacunas. Muchos productos microbianos se han empleado como adyuvantes,1 incluyendo sustancias de Mycobacterium tuberculosis, bacilo de Calmette-Guérin (BCG), Corynebacterium parvum, Brucella abortus y Bordetella pertussis, así como toxoides de Vibrio cholerae y Clostridium tetani. Se han derivado adyuvantes del virus vaccinia y de otros poxvirus, BCG y Salmonella, cada uno transfectado con genes para un antígeno de interés. Otros son derivados de lipopolisacáridos, como el lípido monofosforil A. El adyuvante completo de Freund, que consiste en micobacterias muertas en aceite emulsificado con un antígeno en solución acuosa, se usa en la actualidad con poca frecuencia porque en general causa una intensa reacción inflamatoria local. Otros adyuvantes incluyen extractos de jabón del árbol de tejo (Quillaja saponaria), polímeros (v.gr., inulina), péptidos complejos y diversas citocinas. Por ejemplo, la interleucina-12 (IL-12) parece actuar como un promotor intenso de las reacciones inmunológicas mediadas por células en los ratones.2 La sal de aluminio alumbre también está autorizada para usarse como adyuvante en humanos.
Los anticuerpos son un grupo heterogéneo de proteínas séricas denominadas inmunoglobulinas. Las inmunoglobulinas son secretadas por células B diferenciadas denominadas células plasmáticas. De acuerdo con la teoría de Sir Francis MacFarlane Burnett de la selección clonal, una sola célula plasmática produce un solo anticuerpo específico. Esta teoría se ilustra en forma clara en el mieloma múltiple, en el que ocurre proliferación maligna de células plasmáticas en la médula ósea. La célula plasmática del mieloma múltiple produce una inmunoglobulina monoclonal anormal denomina proteína del mieloma. Su homogeneidad es claramente visible en la electroforesis, en la que se forma una sola banda densa. Ciertos anticuerpos que se producen en respuesta a antígenos muy homogéneos, como el polisacárido estreptocócico, pueden ser también homogéneos.
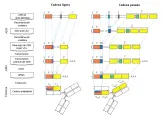
Los monómeros de inmunoglobulina están formados por dos cadenas pesadas y dos cadenas ligeras que son idénticas [ver figura 1a]. Cada cadena ligera está unida a una cadena pesada por puentes disulfuro (S-S) y las cadenas pesadas también están unidas entre sí por uno o más puentes S-S. Las secuencias de aminoácidos en las cadenas pesadas y ligeras están divididas en regiones que son constantes o variables [ver figura 1b]; además, cada región variable contiene secuencias que son hipervariables.
|
|
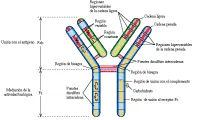

CLASIFICACION DE LAS INMUNOGLOBULINAS
Existen cinco clases principales de inmunoglobulinas y cada clase está compuesta de una cadena pesada específica: la IgG tiene dos cadenas g; la IgM, dos cadenas µ; la IgA, dos cadenas a; la IgD, dos cadenas d y la IgE, dos cadenas e [ver tabla 1]. Así mismo, existen dos tipos de cadenas ligeras, k y l, las cuales pueden diferenciarse por medio de antígenos. La forma secretada de IgM es un pentámero compuesto de cinco monómeros [ver figura 2]; la IgM también puede existir como monómero unido a superficies. La IgA se encuentra en dos formas, como monómero o como dímero. Las inmunoglobulinas poliméricas, IgM e IgA, tienen una cadena J adicional (de unión), que ayuda en su polimerización.
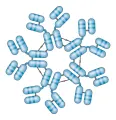
|
| Figura 2 |
| Forma pentamérica de la IgM. |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
IgG La IgG es la inmunoglobulina principal en el suero, donde se
encuentra en forma de monómero. Tiene una vida media en sangre cercana a
23 días y es el principal anticuerpo que se eleva después de un
reto antigénico. Existen cuatro subclases de IgG (IgG1, IgG2, IgG3 e
IgG4), cada una con estructura y propiedades biológicas diferentes. Por
ejemplo, solo la IgG1 y la IgG3 fijan al primer componente del complemento y se
adhieren a los monocitos. Los anticuerpos que cubren a los microrganismos
haciéndolos más susceptibles a la fagocitosis (i.e.,
opsonización) son de la clase IgG. Los anticuerpos IgG también
neutralizan toxinas, como la toxina diftérica, y pueden neutralizar
virus. Aunque el feto no produce esta clase de inmunoglobulina, la IgG cruza
con facilidad la placenta, por lo que los anticuerpos de IgG que se encuentran
en el recién nacido provienen de la madre.
Clínicamente la IgG se ha empleado con éxito para reconstituir la inmunidad de enfermos con agammaglobulinemia y para prevenir la enfermedad hemolítica del recién nacido. Las mujeres con grupo sanguíneo Rh negativo que tienen un feto con eritrocitos Rh1 positivos, pueden ser sensibilizadas durante el primer parto por los eritrocitos Rh1 positivos del feto. Entonces la madre producirá anticuerpos IgG anti-Rh1 que atravesarán la placenta durante embarazos subsecuentes; estos anticuerpos reaccionarán con los eritrocitos fetales, ocasionando la enfermedad hemolítica. La eritroblastosis fetal puede prevenirse inyectando anticuerpos anti-Rh positivos, ricos en IgG (RhoGAM) a la madre Rh1-negativa, durante el parto o poco tiempo después de éste o de un aborto. Los anticuerpos parecen combinarse con cualquier eritrocito fetal Rh1-positivo presente, evitando que éstos inmunicen a la madre.
La IgG puede separarse en tres fragmentos por la enzima proteolítica papaína [ver figura 1a]. Dos de los fragmentos son similares y se denominan Fab, el tercero se llama Fc. La porción Fc es la responsable de la actividad biológica de las diversas inmunoglobulinas. Entre otras cosas, la porción Fc controla la capacidad de las inmunoglobulinas de fijarse a células, fijar complemento y atravesar la placenta. Otra enzima proteolítica, la pepsina, separa la molécula IgG por detrás de los puentes S-S que enlazan las cadenas pesadas, dejando un fragmento grande, F(ab)2, que es capaz de unirse y precipitar al antígeno por su bivalencia y capacidad para formar una red.
IgA La IgA es la inmunoglobulina predominante en las secreciones, donde suele encontrarse en forma de dímero y es liberada como tal por las células plasmáticas que se encuentran en la cercanía. La IgA monomérica constituye el 15 por ciento de las inmunoglobulinas séricas, en el suero tiene una vida media de 5 a 6 días. Existen dos subclases de IgA, IgA1 e IgA2. El dímero se combina con una cadena polipeptídica, denominada pieza secretora, que es producida por las células epiteliales locales. De esta forma, es muy resistente a la digestión proteolítica. A diferencia de la IgA sérica, la IgA combinada con la pieza secretora se transporta en forma activa a través del epitelio mucoso por endocitosis [ver figura 3].
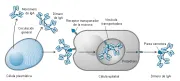
|
| Figura 3 |
| Secreción de IgA |
La IgA está presente en saliva, lágrimas y calostro. También se encuentra en el aparato respiratorio y digestivo, en la vagina y en la próstata. Los mayores niveles de anticuerpos contra antígenos de la dieta en personas con deficiencia de IgA sugieren que este tipo de inmunoglobulina limita en condiciones normales la absorción de dichos antígenos.
Se cree que la IgA juega un papel importante en la inmunidad local al neutralizar virus y al combinarse con virus y bacterias, impidiendo la adherencia de estos microrganismos a la superficie de las mucosas. Aunque la IgA no fija complemento, puede activar a la vía alterna. Uno de los componentes del complemento generado por esta vía, C3b, puede ayudar en la opsonización de las bacterias, aumentando su captación y destrucción por los fagocitos.
IgM La mayoría de las células B tienen IgM monomérica en su superficie. Sin embargo, la IgM existe principalmente como un pentámero que se encuentra en el suero, donde forma el 10 por ciento de las inmunoglobulinas. En la respuesta inmunológica, la IgM es la primera inmunoglobulina en elevarse tras el estímulo antigénico. Las células que producen IgM o sus precursores no se convierten en células de memoria, por lo que una segunda exposición a un antígeno no produce una mayor cantidad de anticuerpos IgM que el primer estímulo. Debido a que la respuesta de IgM contra antígenos es de corta duración, su presencia también puede ser de ayuda para establecer el diagnóstico de una infección en particular. El feto elaborará anticuerpos IgM contra ciertos microrganismos lo que puede ser de ayuda en el diagnóstico de la toxoplasmosis, la rubéola o la sífilis fetal; sin embargo, no todos los fetos infectados por esos microrganismos producen tales anticuerpos.
Como un pentámero, la IgM es muy eficiente para fijar complemento. Molécula por molécula, es 20 veces más efectiva en aglutinar bacterias y eritrocitos y 1,000 veces más activa en las reacciones bactericidas que la IgG. Las isohemaglutininas, como la anti-A y la anti-B, son de la clase IgM, y la macroglobulinemia de Waldenström es un padecimiento que representa la producción monoclonal de IgM.
IgD La IgD, que es un monómero, existe en el suero en cantidades mínimas. Se encuentra en concentraciones relativamente elevadas en la sangre del cordón umbilical. La mayoría de las células B de la sangre del cordón umbilical tienen IgD en su superficie y la mayoría de las células B del adulto tienen, en su superficie tanto IgD como IgM. Se han encontrado células plasmáticas productoras de IgD en las amígdalas y adenoides, aunque su existencia es muy rara en otros tejidos linfoides.
IgE La IgE se encuentra presente en cantidades mínimas en el suero, constituyendo sólo el 0.004 por ciento de las inmunoglobulinas séricas. La IgE es el anticuerpo termolábil (que se conocía en el pasado como reagina) que desempeña una función principal en la hipersensibilidad inmediata, es decir, la reacción inmunológica que se presenta en la fiebre del heno, el asma extrínseco, la urticaria y la anafilaxia. La IgE se une fuertemente a las células cebadas y a los basófilos; cuando estas células cubiertas con IgE interaccionan con antígenos específicos (alergenos) liberan potentes sustancias mediadoras de la hipersensibilidad inmediata, incluyendo a la histamina, la sustancia de reacción lenta de la anafilaxia (SRS-A) y un factor quimiotáctico eosinofílico. Los niveles de IgE son mayores de lo normal en personas con atopia y los antígenos IgE específicos contra un alergeno en particular también están elevados, correlacionando con la susceptibilidad del individuo. En las alergias los anticuerpos IgE específicos se detectan por medio de radioinmunoanálisis (RAST o prueba radioalergosorbente).
La IgE se une a las células cebadas o a los basófilos por medio de su porción Fc. El calor destruye la capacidad de la porción Fc de unirse a la célula. Se encuentran células plasmáticas productoras de IgE en las amígdalas, adenoides y la mucosa de los aparatos respiratorio y gastrointestinal. Receptores precisos para la IgE se localizan sobre la superficie de las células cebadas, las células B, las células T, los macrófagos y los eosinófilos.
Se desconoce la función exacta de la IgE. Ciertamente, las manifestaciones de la hipersensibilidad inmediata, como la fiebre del heno o el asma extrínseco, no parecen tener un propósito útil para el individuo afectado o para la especie en general. Por lo tanto, la observación de que los niveles de IgE se encuentran elevados en ocasiones en personas que viven en las regiones tropicales y en especial en aquéllas afectadas por parásitos helmintos, fue considerada por algunos inmunólogos como una posible indicación de la función de la IgE, es decir, que tiene un papel protector contra los parásitos. Los mediadores liberados podrían afectar a los parásitos, ya sea de manera directa o al producir incremento en la permeabilidad vascular y liberar factor quimiotáctico eosinofílico, lo cual podría causar el acúmulo de otros anticuerpos (v.gr., IgG) y células necesarios para atacar al parásito. En este contexto es de interés el hecho de que los eosinófilos pueden mediar un daño dependiente de IgG contra esquistosómulas (la forma larvaria del parásito Schistosoma mansoni). Además, complejos inmunes de IgE específicos contra parásitos pueden inducir citotoxicidad mediada por macrófagos contra organimos esquistosómulas.
Diferencias antigénicas de las inmunoglobulinas
Existen tres tipos de determinantes serológicos (antigénicos) en las inmunoglobulinas: isotípicos, alotípicos e idiotípicos.
Determinantes isotípicos Los determinantes isotípicos establecen la diferencia entre las regiones constantes de las diversas clases y subclases de cadenas ligeras; representan diferentes genes de la región constante [ver tabla 1]. Por ejemplo, en un individuo existen cuatro isotipos de la cadena pesada de IgG: g1, g2, g3 y g4, que representan las subclases IgG1, IgG2, IgG3 e IgG4, respectivamente. Hay un sólo isotipo de la cadena ligera k y un isotipo de la cadena ligera l.
Determinantes alotípicos Los determinantes alotípicos distinguen entre las inmunoglobulinas de un isotipo en particular, representan alelos diferentes de los genes de las inmunoglobulinas y, por lo tanto, se determinan genéticamente de acuerdo con las leyes mendelianas de manera muy similar a la que determina los grupos sanguíneos ABO. Las moléculas de cadena pesadas g de un individuo pueden tener varios marcadores diferentes, mientras que no se conocen marcadores alotípicos para las cadenas pesadas µ, d y e o para las cadenas ligeras l. Las cadenas pesadas g poseen más de 20 marcadores alotípicos distintos, que se determinan en forma colectiva Gm. Además, las cadenas ligeras k contienen un grupo de tres marcadores alotípicos llamados Km.
Determinantes idiotípicos Un idiotipo se define como un determinante antigénico único localizado en la región variable de un anticuerpo. Un idiotipo es la expresión antigénica de la región variable de un anticuerpo. Los determinantes idiotípicos distinguen a una inmunoglobulina de otra del mismo alotipo.
Origen genético de la diversidad de los anticuerpos
Las mitades carboxi terminales de todas las cadenas ligeras k tienen secuencias de aminoácidos casi idénticas, por lo que esta porción de la molécula se denomina dominio constante o C. La mitad amino terminal tiene una secuencia variable de aminoácidos y se denomina dominio variable o V [ver figura 1]. Los primeros 110 aminoácidos de la porción amino terminal de la cadena ligera l y de la cadena pesada también son variables. El 75 por ciento restante de la cadena pesada es contante y contiene tres regiones homólogas.
Dentro de las regiones variables, tres áreas, conocidas como regiones hipervariables o determinantes de la complementaridad, muestran una variación aún mayor; estas áreas corresponden al sitio de unión entre el antígeno y el anticuerpo [ver figura 4]. El análisis de rayos X ha demostrado que las moléculas de inmunoglobulina están formadas de unidades globulares compactas conectadas por segmentos cortos de cadenas polipeptídicas más o menos lineales [ver figura 1]. Como es de esperarse, las regiones hipervariables se localizan en la interfase entre la inmunoglobulina y el antígeno.
|
|

El aspecto más intrigante del control genético de la síntesis de inmunoglobulinas es la diversidad del producto: las células plasmáticas pueden producir anticuerpos que reaccionan con un número indefinido de sitios antigénicos diferentes. ¿Cómo puede el ADN codificar contra un número tan grande de antígenos, muchos de los cuáles existen (en la escala de la evolución) desde hace muy poco tiempo?
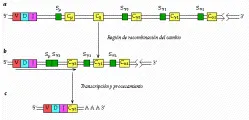
En todas las células, el ADN de la cadena ligera k codifica más de 300 regiones variables (V), cinco regiones de unión (J) y una región constante (C). Las regiones V y J se encuentran separadas del segmento C por una porción de ADN. Por lo tanto, el ADN embrionario tiene la capacidad para formar al menos 1,500 combinaciones diferentes de V y J; en otras palabras, por lo menos 1,500 cadenas ligeras k diferentes.
La aparición de estas líneas celulares individuales es el resultado de la recombinación somática en el ADN y de la unión del ARN [ver figura 5]. Al diferenciarse las células pre-B en células plasmáticas, deleciones en el ADN hacen que uno de los genes V, escogidos al azar, sea adyacente a uno de los genes J. Esta unidad V-J y el resto de las regiones J están separadas del gen C por una porción corta de ADN. En el siguiente paso, el ADN es trancrito al ARN nuclear y la secuencia queda lista para un segundo rearreglo.
|
| Figura 5 |
| Organización genética de las regiones variables |
Este evento se inicia cuando una enzima rompe el ARN nuclear para producir ARN mensajero (ARNm). En un proceso denominado unión del ARN, el segmento que separa a la región o unidad V-J de la región C es removido junto con los segmentos J sobrantes. El segmento V-J-C restante se traduce en una de las 1,500 o más proteínas de las cadenas ligeras k. En realidad el número es mayor por el hecho de que la unión de cualquier porción V con J puede afectar alguna entre una variedad de secuencias de bases apareadas en el sitio de recombinación.
La variabilidad en la cadena pesada también contribuye de manera importante a la especificidad de un anticuerpo. Esta variabilidad es consecuencia de un proceso que incluye la recombinación somática y la unión de ARN, similares al proceso que crea la variabilidad en las cadenas ligeras. El ADN embrionario lleva las instrucciones para la formación de varios cientos de genes V de cadena pesada, seis genes J, 10 a 20 regiones D (diversidad) y nueve genes C (estos genes C codifican las clases de cadenas pesadas: IgM, IgD, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2 e IgE). La deleción de ADN, su transcripción al ARN nuclear y la unión del ARN producen la secuencia V-D-J-C final en el ARNm, que es codificada por los ribosomas a una proteína de cadena pesada. Este proceso de ensamble produce más de 18,000 variedades posibles en las proteínas de las cadenas pesadas (la especificidad del anticuerpo no varía con la clase del mismo, por lo que los nueve genes C no entran en el cálculo).
La combinación de las más de 1,500 variedades de cadenas ligeras con las 18,000 variedades de cadenas pesadas podría producir más de 27 millones de anticuerpos diferentes con sitios de combinación para antígenos diferentes. Además, ocurre mutación somática, en especial durante la maduración de la afinidad y la velocidad de la mutación somática es relativamente alta (un par de bases por 1,000 divisiones celulares). Por lo tanto, el número potencial de anticuerpos específicos en una persona es indefinido.
Aunque algunos de los mecanismos del rearreglo V-D-J son característicos de las células B (y de las células T porque la síntesis de los RCT ocurre por un mecanismo semejante [ver adelante, Receptores de las células T]), también participan los mecanismos generales de reparación del ADN.3 Dos de los genes incluidos en el rearreglo V-D-J en las células T y B son los genes activadores de la recombinación rag1 y rag2.4-6 La ruptura de estos genes causa un bloqueo en el desarrollo de las células B antes de la transición de células pro-B a pre-B, como se observa en los pacientes con síndrome de inmunodeficiencia combinada severa o de Omenn.7 Del mismo modo, la alteración del gen de la IgM de superficie, de la región J de la cadena pesada o de la región J de la cadena ligera k causa un bloqueo parecido en el desarrollo de las células B.
En cada célula B individual solo uno de los cromosomas sufre recombinación V-D-J completa, causando la expresión de cadenas pesadas y ligeras. Existe un mecanismo que evita que el otro cromosomas se rearregle y por lo tanto se exprese en la misma célula. A esto se denomina exclusión alélica. Este proceso evita que una célula B exprese dos inmunoglobulinas o RCB totalmente diferentes. En las células T opera un mecanismo semejante durante la síntesis del RCT [ver adelante, Receptores de células T].
Respuestas de anticuerpos primaria y secundaria
Cuando un antígeno se introduce por primera vez en el organismo ocurre una respuesta primaria que se caracteriza por una fase de latencia que dura varios días y durante la que no se detectan anticuerpos. Aparecen cantidades cada vez mayores del anticuerpo IgM en el suero, que suelen alcanzar su máximo después de 7 días. Después de 6 o 7 días se detecta también anticuerpo IgG. El título de IgM comienza a disminuir antes de que se alcance el título máximo de IgG, alrededor de 10 a 14 días después de que se introdujo el antígeno. Después disminuye el título de anticuerpo y 4 o 5 semanas después de la administración de una sola dosis de antígeno se detecta muy poco anticuerpo.
Si el antígeno se introduce por segunda ocasión ocurre una respuesta secundaria (también denominada anamnésica o de refuerzo) por la existencia de células B de memoria. En este caso los títulos tanto de IgM como de IgG aumentan en forma exponencial, si la fase de latencia observada en la respuesta primaria. Mientras que el nivel máximo de IgM puede ser igual o un poco mayor que el observado en la respuesta primaria, el nivel máximo de IgG durante la respuesta secundaria es mucho mayor y dura más. Esta variación en la respuesta es una ilustración obvia de la memoria inmunológica y es causada por proliferación de las células B específicas de antígeno y de las células T cooperadoras durante la respuesta primaria. Las características de las respuestas primaria y secundaria explican la necesidad de administrar dosis de refuerzo en los programas de inmunización.
Las propiedades de unión del anticuerpo cambian con el tiempo. A este proceso se le denomina maduración de la afinidad, e incluye el proceso de maduración somática y de selección. Después del estímulo primario, los anticuerpos tienen una afinidad cada vez mayor por el antígeno según va pasando el tiempo, y forman complejos antígeno-anticuerpo con una mayor estabilidad. Además, los anticuerpos se vuelven menos específicos y presentan más reacciones cruzadas con antígenos relacionados. En realidad, la disminución en la especificidad refleja el hecho de que las reacciones cruzadas, que previamente eran muy débiles para ser detectadas, se vuelven aparentes al tener el anticuerpo mayor afinidad por el antígeno.
CAMBIO DE CLASE DE LA INMUNOGLOBULINA
Los genes que codifican para las cadenas pesadas de IgM e IgD (genes µ y d, respectivamente), tienen un papel crucial en la respuesta inmune primaria. Mientras que los anticuerpos de IgM son incapaces de actuar en muchos tejidos del organismo, la IgG y la IgA tienen funciones en el sistema inmunológico periférico. El cambio de clase significa que la misma región variable puede transferirse de la cadena pesada de una IgM a una de otro anticuerpo. En el cambio de producción de IgM a IgG que constituye la respuesta secundaria, genes de la región constante sufren deleción antes de que el ADN se transcriba al ARN [ver figura 6].7,8 Si la célula cambia para producir IgG3, por ejemplo, los genes para m y d sufren deleción [ver figura 7]. Después de la transcripción, la unión del ARN produce un ARNm con la secuencia V-D-J-Cg3, que se traduce a la proteína.
|
|
Receptores de inmunoglobulinas
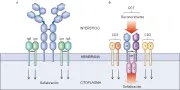
Además de secretarse, las inmunoglobulinas pueden expresarse en la superficie de las células B, en donde actúan como receptores de antígenos.9 Estas inmunoglobulinas de superficie de membrana (IgSm) difieren de las inmunoglobulinas secretadas en que tienen un dominio transmembrana y son monoméricas. La primera IgSm que expresa una célula B es IgM, en una fase más avanzada del desarrollo se expresa en forma conjunta IgD. La IgSm no viaja a la superficie celular por sí misma, sino que el proceso requiere la formación de un complejo consistente en la Ig y dos cadenas polipeptídicas denominadas Iga e Igb en el retículo endoplásmico [ver figura 8a]. El RCB resultante se une al antígeno y esto provoca la maduración de la célula B. La estimulación de la célula T cooperadora activa a la célula B, haciendo que se diferencie a una célula de memoria o plasmática, que produce anticuerpos secretorios específicos para el antígeno encontrado. Después de la diferenciación terminal no se expresan más Iga o Igb. La célula plasmática madura cesa de expresar IgSm, a pesar de que mantenga el ARNm para ésta.
|
| Figura 8 |
| Receptores de la célula B y T |
El RCB tiene un papel importante en la regulación de la respuesta inmunológica. La respuesta de la célula B al antígeno puede volverse anérgica, proporcionando un mecanismo de control para las respuestas de las células B y la producción de anticuerpos. Se piensa que un precursor del RCB que se expresa en la superficie de las células pre-B controla la conclusión alélica. Además, los RCB interactúan con receptores Fc.
Los receptores para Fc fijan la porción Fc de una inmunoglobulina, se expresan en múltiples células, incluyendo células cebadas, macrófagos, eosinófilos y células tumorales. Los receptores para Fc constituyen una familia de moléculas. El receptor de Fc para la IgE (FceRI) sirve como modelo para todos los receptores de este tipo, y consiste en tres cadenas polipeptídicas, designadas como a, b, y g. El FceRI media la transducción de señales en la célula cebada cuando la IgE se une al receptor. La cadena a es el sitio de unión para la porción Fc de la IgE. La cadena b es una molécula transmembrana que conecta a FceRI a con FceRI g, la cadena responsable de reclutar las moléculas encargadas de las señales de transducción.10,11 El FcRIIbI, otro receptor Fc se expresa en las células B y proporciona una señal de retroalimentación negativa al RCB, lo que produce la terminación de las respuestas inmunes humorales.12
Anticuerpos creados por ingeniería
El desarrollo del hibridoma de linfocitos, productos de fusión celular, tiene implicaciones revolucionarias para la inmunología y la medicina clínica. Los hibridomas de linfocitos constituyen el medio por el cual pueden producirse títulos muy altos de anticuerpos puros, muy específicos, para fines experimentales. El hibridoma de linfocitos, tal y como fue desarrollado por Köhler y Milstein,13 es el producto de la fusión de una célula de mieloma de ratón y un linfocito obtenido del bazo de un ratón inmunizado con un antígeno específico. Los hibridomas pueden ser clonados y seleccionados para la producción específica anticuerpos.
ANTICUERPOS MONOCLONALES HUMANOS
En la actualidad existen varios métodos para elaborar anticuerpos monoclonales humanos. El primero consiste en tomar el ADN complementario que codifica un anticuerpo monoclonal murino y remplazar en forma sistemática las secuencias murinas con secuencias humanas. El ADN que codifica para las regiones constantes de las cadenas pesadas y ligeras humanas remplaza las secuencias de ADN murinas de las regiones variables con humanas por medio de mutagénesis, dejando las regiones hipervariables intactas. Una desventaja de este método es que puede afectar la afinidad.
Otro método de humanizar anticuerpos monoclonales murinos consiste en crear un ratón transgénico que contenga grandes segmentos de ADN humano, incluyendo varias regiones variables y todas las regiones constantes humanas.14 Debido a que este ratón tiene aún sus propias regiones de inmunoglobulinas, se cruza con un ratón en el que exista ruptura dirigida de la región J de los genes de las cadenas pesadas de Ig y ligeras k. La progenie de esta cruza puede inyectarse con cualquier antígeno y se producirán anticuerpos monoclonales humanizados. Debido a que el ratón transgénico contiene un número limitado de regiones variables humanas, la potencia de estos anticuerpos depende de la mutación somática natural y la maduración de la afinidad que ocurra en el ratón.
Un tercer método de generar anticuerpos monoclonales humanos consiste en construir bibliotecas (inventarios) de regiones variables humanas en bacterias o en bacteriófagos. En teoría, pueden clonarse todas las regiones variables. El anticuerpo puede expresarse en la superficie de la bacteria o el bacteriófago y seleccionarse por afinidad al antígeno de interés.
Los anticuerpos monoclonales se están empleando en muchos tratamientos. Por ejemplo, se han empleado con éxito anticuerpos monoclonales humanizados contra factor de necrosis tumoral-a (FNT-a) en el tratamiento de la enfermedad de Crohn. En otras aplicaciones, se emplean anticuerpos monoclonales para eliminar células T y células tumorales antes del trasplante de médula ósea y durante el rechazo agudo de trasplantes. Otros usos potenciales de los anticuerpos monoclonales son la producción de anticuerpos anti-IgE y receptores anti-hormonas para prevenir alergias y modular alteraciones endócrinas, respectivamente.
A diferencia de los receptores de inmunoglobulinas de las células B, que pueden reconocer antígenos libres, los receptores de células T sólo reconocen antígenos combinados con antígenos autólogos del complejo principal de histocompatibilidad, por lo general sobre la superficie de los macrófagos, aunque pueden estar también sobre la superficie de una célula B, una célula dendrítica o una célula endotelial. Las células T cooperadoras CD4+ (así como las pocas células T citotóxicas CD4+), requieren moléculas clase II del CPH, y las células T citotóxicas CD8+ requieren moléculas clase I del CPH. Este fenómeno se conoce como restricción del CPH. La capacidad de las células T para reconocer estas moléculas del CPH propias se establece durante el desarrollo en el timo antes de que los linfocitos se expongan al antígeno.
Las moléculas que forman el RCT y los genes que codifican estas moléculas han sido ya aisladas y clonadas. El RCT está compuesto de seis polipéptidos distintos conocidos como complejo CD3-RC T.15-17 Del 85 al 95 por ciento de los linfocitos de la sangre periférica tienen receptores RCT-ab y sólo el 5 al 15 por ciento tienen RCT-gd. La porción del complejo RCT-ab que reconoce al antígeno está compuesta de dos cadenas polipeptídicas glucosiladas, denominadas RCT-a y RCT-b, que están unidas por puentes disulfuro, formando un heterodímero. Las cadenas polipeptídicas correspondientes en el complejo RCT-gd consisten en RCT-g y RCT-d. Cada una de las cadenas RCT-a, RCT-b, RCT-g y RCT-d contienen porciones variables (V) y constantes (C) análogas a las observadas en las inmunoglobulinas.
Los heterodímeros de RCT-ab y RCT-gd están muy relacionados con las proteínas del CD3: CD3-g, CD3-d, Cd3-e y CD3-s [ver figura 8b]. Las proteínas CD3 están presentes en todas las células T de sangre periférica y en el 90 por ciento de los timocitos. Las expresiones del CD3 y de los complejos RCT sobre la superficie celular son mutuamente dependientes: no se observa ninguno de estos complejos en la superficie celular sin el otro. Datos estructurales y funcionales sugieren que las actividades del RCT se distribuyen entre las subunidades del complejo RCT-CD3: los polipéptidos RCT (a, b, g y d) se fijan al antígeno y productos del CPH y las proteínas CD3 transfieren la señal al citoplasma de la célula T lo que activa sus funciones.
Al usar ratones que contienen mutaciones espontáneas o creadas de RCT se ha confirmado la vía de desarrollo de la célula T [ver figura 9]. La organización de los genes que codifican el RCT-a, -b, -g y -d es análogo al de los genes de las cadenas pesadas de las inmunoglobulinas: existen segmentos V, D y J, que están limitados por secuencias de reconocimiento que median la recombinación local específica [ver antes, Anticuerpos]. Por lo tanto, la diversidad de os RCT se genera por muchos de los mismos mecanismos que son usados por las células B para la producción de las imunoglobulinas. De hecho, es posible que las células T y B usen la misma enzima de recombinación, la recombinasa.
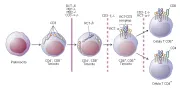
|
| Figura 9 |
| Vía de desarrollo de una célula T |
Las secuencias genómicas que codifican la cadena b del RCT contienen dos genes muy semejantes en la región constante, Cb1 y Cb2, cada uno de los cuales se asocia con un grupo de 6 o 7 genes J y un solo gen D. Existen por lo menos 70 genes Vb que se asocian con los dos genes Cb. Estas regiones variables son diferentes a las regiones variables de las inmunoglobulinas. El rearreglo de los segmentos del gen de la cadena b puede permitir la producción de alrededor de 3,600 cadenas b diferentes.
Los genes de las cadenas a del RCT se rearreglan diferente. Un solo gen Ca es precedido por una tira muy grande de ADN que contiene por los menos 50 genes J diferentes. No se ha demostrado en forma directa un gen Da. Algunos genes Va se organizan como familias de genes relacionados. El rearreglo de los segmentos genéticos de la cadena a puede causar alrededor de 2,500 polipéptidos diferentes. No se han detectado mutaciones somáticas en los RCT. Por lo tanto, pueden formarse alrededor de 107 RCT-ab. Los genes para CD3g, CD3d, CD3e y CD3s se transcriben en todas las células; sin embargo, no sufren rearreglos.
Figura 1a Talar Agasyan
Figura 1b Dana Burns-Pizer. Modelo gráfico de computadora por A. J. Olson, Ph.D. c 1985 Research Institute of Scripps Clinic. Usado con autorización.
Figuras 2, 3, 6, 8 y 9 Seward Hung.
Figura 4 Albert Miller.
Figura 5 Seward Hung.
Figura 7 Dana Burns-Pizer.
Bibliografía
- Furlong ST, David JR: Adjuvants. Therapeutic Immunology. Austen KF, Burakoff SJ, Rosen FS, Eds. Blackwell Scientific Publications, Cambridge, 1996
- Afonso LC, Scharton TM, Vieira LQ, et al: The adjuvant effect of interleukin-12 in a vaccine against Leishmania major. Science 263:235, 1994 [PMID 7904381 ]
- Frank KM, Sekiguchi JM, Seidl KJ, et al: Late embryonic lethality and impaired V(D)J recombination in mice lacking DNA ligase IV. Nature 396:173, 1998 [PMID 9823897 ]
- Oettinger MA, Schatz DG, Gorka C, et al: RAG-1 and RAG-2, adjacent genes that synergistically activate V(D)J recombination. Science 248:1517, 1990 [PMID 2360047 ]
- Hofker MH, Walter MA, Cox DW: Complete physical map of the human immunoglobulin heavy chain constant region gene complex. Proc Natl Acad Sci USA 86:5567, 1989 [PMID 2501792 ]
- Seidl KJ, Manis JP, Bottaro A, et al: Position-dependent inhibition of class-switch recombination by PGK-neor cassettes inserted into the immunoglobulin heavy chain constant region locus. Proc Natl Acad Sci USA 96:3000, 1999 [PMID 10077626 ]
- Villa A, Santagata S, Bozzi F, et al: Partial V(D)J recombination activity leads to Omenn syndrome. Cell 93:885, 1998 [PMID 9630231 ]
- Rolink A, Melchers F, Andersson J: The SCID but not the RAG-2 gene product is required for S mu-S epsilon heavy chain class switching. Immunity 5:319, 1996 [PMID 8885865 ]
- Kurosaki T: Genetic analysis of B cell antigen receptor signaling. Annu Rev Immunol 17:555, 1999
- Kinet JP: The high-affinity IgE receptor (Fc epsilon RI): from physiology to pathology. Annu Rev Immunol 17:931, 1999
- Metzger H: It's spring, and thoughts turn to...allergies. Cell 97:287, 1999
- Scharenberg AM, Kinet JP: PtdIns-3,4,5-P3: a regulatory nexus between tyrosine kinases and sustained calcium signals. Cell 94:5, 1998 [PMID 9674420 ]
- Köhler G, Milstein C: Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 256:495, 1975
- Lonberg N, Taylor LD, Harding FA, et al: Antigen-specific human antibodies from mice comprising four distinct genetic modifications. Nature 368:856, 1994 [PMID 8159246 ]
- Terhorst C, Spits H, Staal F: T lymphocyte signal transduction. Molecular Immunology. Hames BD, Glover DM, Eds. IRL Press, Washington, DC, 1995, p 237
- Garcia KC, Teyton L, Wilson IA: Structural basis of T cell recognition. Annu Rev Immunol 17:369, 1999 [PMID 10358763 ]
- Davis MM, Boniface JJ, Reich Z, et al: Ligand recognition by alpha beta T cell receptors. Annu Rev Immunol 16:523, 1998 [PMID 9597140 ]
- Tonegawa S: The molecules of the immune system. Sci Am 253(4):122, 1985
- Gally JA: Structure of Immunoglobulins. The Antigens. Sela M, Ed. Academic Press, New York, 1973
- Frangione B: Immunogenetics and Immunodeficiency, 2nd ed. Medical and Technical Publishing Co, London, 1975
- Kabat EA: Structural Concepts in Immunology and Immunochemistry, 2nd ed. Holt, Rinehart & Winston, New York, 1976
- Underdown BJ, Schiff JM: Immunoglobulin A: strategic defense initiative at the mucosal surface. Annu Rev Immunol 4:389, 1986 [PMID 3518747 ]
- Cohn M, Blomberg B, Geckeler W, et al: First order considerations analyzing the generator of diversity. The Immune System: Genes, Receptors, Signals. Sercarz EE, Williamson AR, Fox CF, Eds. Academic Press, New York, 1974
- Kabat EA, Bilovsky H, Wu TT: Data Compiled by the Prophet Computer System of the National Institutes of Health (unpublished)
- Wu TT, Kabat EA: An analysis of the sequences of Bence Jones proteins and myeloma light chains and their implications for antibody complementarity. J Exp Med 132:211, 1970 [PMID 5508247 ]
- Janeway CA Jr, Travers P: Immunology. The Immune System in Health and Disease, 3rd ed. Current Biology Ltd, London, 1997
- Cooper M, Lawton AR III: The development of the immune system. Sci Am 231(5):50, 1974
- Mombaerts P, Iacomini J, Johnson RS, et al: RAG-1-deficient mice have no mature B and T lymphocytes. Cell 68:869, 1992 [PMID 1547488 ]