Contenido del artículo
IV SISTEMA DEL COMPLEMENTO
DR. DOUGLAS FEARON
DR. JOHN DAVID
Componentes del sistema del complemento
El sistema del complemento, compuesto por 18 proteínas plasmáticas, es el principal efector humoral de la inflamación inducida por mecanismos inmunológicos [ver tabla 1]. Desempeña un papel fundamental en la resistencia a infecciones, tanto mediadas inmunológica-mente como inespecíficas, así como en la patogenia de lesiones tisulares. Los productos de la activación del complemento regulan varios eventos biológicos, incluyendo la liberación de mediadores químicos por parte de las células cebadas, que aumentan la permeabilidad vascular. Los factores activados del complemento promueven también la contracción del músculo liso y la quimiotaxis de neutrófilos, células mononucleares y eosinófilos, además de favorecer la fagocitosis por adherencia inmune. Entre otros efectos de la activación del complemento se encuentran la solubilización de complejos inmunes, la lisis de membranas celulares, la neutralización de virus y la destrucción de ciertas bacterias.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
La activación del complemento se puede iniciar por cualquiera de dos vías: la clásica o la alterna. Aunque ambas vías culminan con la formación de enzimas que activan el mismo componente crucial, C3, al parecer cada vía tiene una función distinta en la protección del organismo, ya sea contra enfermedades autoinmunes o contra infecciones. Los individuos con deficiencias genéticas de los componentes de la vía clásica están predispuestos a las enfermedades moduladas por complejos inmunes, mientras que las personas con deficiencias de los componentes de la vía alterna sufren ciertas infecciones bacterianas. La función de las dos vías se relaciona con los mecanismos por los cuales se activan: la vía clásica se activa por anticuerpos en forma de complejos antígeno-anticuerpo, mientras que la vía alterna se activa por bacterias en presencia, e incluso en ausencia, de anticuerpos.
La nomenclatura del sistema del complemento es compleja. A los componentes del complemento activados por la vía clásica, es decir por anticuerpos, así como a aquéllos que constituyen la vía lítica terminal, que lisa las células, se les denomina con las claves C1 a C9. A los componentes de la vía alterna, o de la properdina, se les asignan letras; de esta manera, la properdina se denomina P y el factor B se denomina simplemente B. La colocación de una barra sobre la clave de un componente, por ejemplo, C-1 indica que dicho componente ha sido activado. A los productos de la fragmentación de un componente se les asignan letras minúsculas junto con la clave original; por ejemplo, C3 C3a + C3b. Cuando un componente o fragmento ha sido inactivado, se le añade la letra i al principio o al final de su clave, como en iC3b o Bbi.
Los componentes de las vías clásica y alterna participan en una cascada de reacciones proteolíticas limitadas. Típicamente, la separación de una unión polipeptídica en una molécula precursora inactiva libera un fragmento menor, que puede tener actividad biológica propia, y un fragmento mayor. El fragmento mayor puede participar de nuevo en una reacción proteolítica o puede unirse y modificar la especificidad de una proteasa previamente formada, lo cual conduciría a la fragmentación de la siguiente proteína en la cascada. Algunas de las reacciones que pertenecen a la secuencia lítica terminal entrañan la formación de complejos proteína-proteína que no da como consecuencia la separación de uniones polipeptídicas.
Tanto la vía clásica como la alterna provocan la fragmentación de C3. Este paso inicia una secuencia terminal común que genera la mayor parte de las actividades biológicas del sistema del complemento. Aunado a ello, algunos componentes de la vía alterna desempeñan un papel importante en la amplificación de la actividad del complemento [ver figura 2].
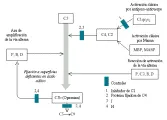
|
| Figura 2 |
| Ruptura de C3: vía clásica y alterna del complemento |
Una vez que el sistema del complemento ha sido activado, es controlado por cierto número de proteínas. Además, las enzimas que fragmentan a C3 y C5, denominadas convertasa de C3 de C5, disminuyen rápidamente.
La vía clásica
La vía clásica del complemento puede ser activada por IgM, IgG1, IgG2 e IgG3 unidas a antígenos. Debido a que la IgM es un pentámero (por lo que tiene cinco sitios de unión por molécula), una sola molécula de IgM es suficiente para fijarse al primer componente del complemento, C1. Por el contrario, se necesitan dos moléculas adyacentes de IgG para establecer la misma unión con C1. Por tanto, la activación por IgG es menos eficiente que por IgM.
C1 está compuesto por tres proteínas, o subcomponentes, denominados C1q, C1r2 C1s2. El factor C1q está formado por 18 cadenas polipeptídicas, mientras que las otras dos proteínas son dímeros polipeptídicos. Cada subcomponente posee una función única: C1q se une a la inmunoglobulina, C1r2 activa a C1s2 y C1s2 activa a C4 [ver figuras 3a a 3e]. La estructura de Clq semeja la de seis tulipanes entrelazados por sus tallos [ver figura 4]. Las estructuras en forma de flor son las porciones globulares de la molécula, que se unen a la región Fc de la IgM o de la IgG y le dan a C1q una valencia potencial de seis. Las estructuras en forma de tallo recuerdan a la colágena y proporcionan los sitios de interacción de C1q con C1r2 y C1s2. C1r2 y C1s2 son proteasas de serina que se convierten de sus precursores a sus formas activas por reacciones proteolíticas. C1r2 se activa en forma autocatalítica y C1s2 se activa por C1r2-. La activación autocatalítica de C1r2 suele evitarse por el inhibidor de C1 (C1-INH). Sin embargo, cuando C1q se fija a una inmunoglobulina que está unida al antígeno, se evita el acceso de C1-INH a C1r2, ocurriendo la autoactivación. C1s2 también está sujeta a inhibición por C1-INH, lo cual limita el número de moléculas de C4 y C2 que este subcomponente puede activar [ver figura 2].
|
|
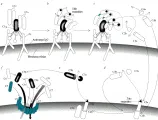
La convertasa C42 actúa fragmentando a C3 en dos partes [ver figura 3f]. El fragmento menor (C3a), que es liberado en la fase líquida, es un péptido que puede liberar mediadores de las células cebadas, siendo muy activo en concentraciones bajas. El otro fragmento (C3b) tiene la capacidad de establecer uniones covalentes con las membranas celulares. Si se une por contiguidad a C42, modifica la especificidad de la enzima, permitiéndole actuar sobre C5. Al C42 modificado por C3b se le denomina convertasa de C5. Además de lo anterior, C3b adherido a la membrana celular induce inmuno-adherencia, es decir, facilita la unión de una célula cubierta de C3b a fagocitos con receptores para dicha proteína, con lo cual estimula la fagocitosis. La C3b es inactivada por I en presencia de H (antes llamado ß1H).
La secuencia terminal
La convertasa de C5, C423b, divide a C5 en un fragmento pequeño, C5a ,y uno mayor, C5b. C5a tiene propiedades similares a las de C3a, además de que es quimiotáctico para leucocitos. C5b se une a C6; la fracción C5b6 formada de este modo puede unirse a C7, ya sea mientras está unida a la convertasa de C5 o en la fase líquida. El complejo C5b67 puede separarse de la convertasa de C5 y unirse a la membrana celular en otro punto. Además, se puede unir a membranas de células no sensibilizadas, en un proceso conocido como lisis reactiva, lo cual extiende los efectos citolíticos del complemento a células llamadas observadoras inocentes. Cuando C5b67 no se une a una membrana celular, se convierte en C5i67, molécula litícamente inactiva.
Cuando el complejo C5b67 entra a la membrana, se une a una molécula de C8, produciendo daño a la membrana y permitiendo el flujo de iones hacia el interior. Este nuevo complejo puede unirse hasta a seis moléculas de C9, proceso que induce una lisis rápida. Si esta secuencia se efectúa en una bacteria gram negativa, ésta será destruida por el complejo de ataque terminal.
La cadena de eventos desde la fijación de C1q hasta la fijación de C9 y la lisis de la célula entraña la acción de muchas proteínas, pero de solo dos nuevos complejos enzimáticos, C1 y C4b2a. Este último se modifica en presencia de C3b para formar la convertasa de C5. El resto de la secuencia de eventos ocurre mediante interacciones proteína-proteína.
La vía alterna
La incubación en suero fresco con diversas bacterias, hongos, algunos parásitos, células heterólogas de mamífero o complejos inmunes insolubles producirá fragmentación de C3, activando la secuencia terminal del complemento. A este sistema de activación del sistema del complemento se le ha llamado la vía alterna. Dado que esta vía no requiere de anticuerpos, puede tener un papel predominante en la defensa no inmunológica inicial contra infecciones.
Se ha utilizado extensamente el zimosán, un polisacárido insoluble de las levaduras, para estudios de la vía alterna. La adición de zimosán a un suero que contenga Mg2+ produce activación de C3 aún en ausencia de C1, C4 y C2. Se requiere de varias proteínas séricas, incluyendo las proteínas P; B que es una proteína similar a C2; D, enzima que fragmenta a B en Bb; y C3.
En esta vía se forman dos tipos de convertasa de C3, una preliminar y otra amplificadora. La función de la convertasa de C3 preliminar es aportar la pequeña cantidad de C3b necesaria para producir la convertasa amplificadora. La convertasa preliminar se genera a partir del componente C3. El C3 contiene un tioéster interno entre los residuos de glutamil y cisteinil que es sometido lenta y espontáneamente a hidrólisis. Aquellas moléculas de C3 cuyo tioéster ha sido hidrolizado adquieren una estructura semejante a la del C3b y pueden unirse a B en forma reversible en presencia de Mg.2+ El B fijo es separado por D para formar C3, Bb, la convertasa preliminar de C3. Esta enzima separa a las moléculas adicionales de C3 en sus fragmentos C3b y C3a.
En presencia de Mg,2+ C3b se une a B (en forma similar a la unión de C4b con C2), formando C3b, B [ver figura 6]. D separa una porción de B para formar C3b,Bb, la conver-tasa amplificadora de C3. Esta enzima es capaz de fragmentar cantidades mucho mayores de C3 y de activar la secuencia terminal. Conforme C3b, Bb fragmenta a C3, se produce una mayor cantidad de C3b, que a su vez se une a más B. Las moléculas adicionales de C3b,Bb formadas de este modo pueden fragmentar más C3, amplificando la reacción. P aumenta la reacción al unirse y estabilizar a C3b,Bb. Las moléculas adicionales de C3b producidas por la fragmentación de C3 se unen también a la membrana celular en sitios adyacentes a C3b,Bb, con lo cual le confieren a esta molécula especificidad para C5, lo cual permite la activación de la secuencia terminal del complemento.
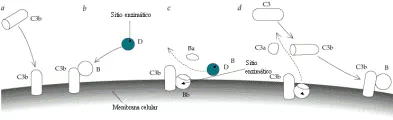
|
| Figura 6 |
| Vía alterna de activación del complemento |
La acción de dos proteínas reguladoras, H e I, hace posible que no toda la C3b generada por la fragmentación preliminar de C3 pase a formar parte de la convertasa amplificadora y automáticamente fragmente una mayor cantidad de dicho componente. H se une a C3b, bloqueando la captación de B e incluso produciendo la disociación de B o Bb que ya se encuentre unida a C3b. De esta manera, H bloquea la formación y funcionamiento de la convertasa de C3 amplificadora. I es una proteasa de serina que convierte a C3b en iC3b, forma inactiva de dicha proteína. Para este efecto de I se requiere de la presencia simultánea de H. Es posible que H altere reversiblemente la conformación de C3b y exponga el sitio sobre el que actúa I. La extracción de cualquiera de estas dos proteínas del suero afecta la regulación de la convertasa de C3 amplificadora, conduciendo a la rápida fragmentación de C3 y B.
La activación de la vía alterna del complemento significa que hay un defecto en el control de la convertasa de C3 amplificadora. La unión de C3 a ciertas superficies celulares interfiere con el control de C3b mediado por H. La C3b que se encuentra en la fase líquida o unida a una superficie no activadora de la vía se une a H con una afinidad 100 veces mayor que aquélla con la que se une a B. Dicha C3b unida a H puede entonces ser inactivada por I. Por el contrario, la C3b que está unida la superficie de un activador de la vía alterna, como el zimosán, tiene una afinidad menor por H, lo que hace posible la unión con B para formar C3b,Bb. La convertasa de C3 así formada fragmentará entonces a C3, formando C3b, con lo cual perpetuará la reacción de amplificación.
El ácido siálico de la superficie celular, presente en glucoproteínas, glucolípidos y polisacáridos, aumenta la afinidad por H de la C3b unida a esa superficie celular. Por lo tanto, las células que contienen cantidades abundantes de ácido siálico no activan la vía alterna del complemento, mientras que aquellas células que carecen de dicho carbohidrato son activadoras. La mayor parte de las bacterias y todos los vegetales carecen de ácido siálico. Por lo tanto, la capacidad de la vía alterna del complemento para responder a células que son deficientes en dicho carbohidrato puede estar relacionada con el papel natural de dicha vía en la resistencia a infecciones. En apoyo a esta teoría tenemos el hecho de que algunas especies bacterianas que sí contienen ácido siálico son más patógenas para los recién nacidos que organismos similares que carecen de dicho carbohidrato; entre las bacterias con ácido siálico en su cápsula se encuentran Streptococcus tipo III del grupo B, Neisseria meningitidis de los grupos B y C y Escherichia coli K1.
Los parásitos humanos como Leishmania major y Trypanosoma cruzi han desarrollado varias estrategias para evadir el reconocimiento por la vía alterna, incluyendo la síntesis de glucoproteínas que contienen ácido siálico, la rápida degradación del factor C3b unido y la síntesis de una proteína que compite con el factor B por la unión a C3b.
Las células de huésped están protegidas de los efectos de la activación del complemento cuando menos por tres proteínas de membrana: el factor acelerado de la degradación (FAD), la proteína cofactor de membrana (PCM) y el CD59. El FAD y la PCM interactúan con C4b y C3b unidos a las células y evitan la activación de la vía clásica y alterna. El CD59 inhibe la unión de C9 al complejo C5b678, con lo cual evita la formación del complejo de ataque C5b-C9.
Tanto el FAD como el CD59 se fijan en la bicapa lípida por uniones glucolípidas. Esta estructura facilita la rápida difusión lateral de estas proteínas y con ello permite a las moléculas individuales la protección de grandes áreas de la membrana. Una deficiencia en estas dos proteínas causa la mayor sensibilidad de los eritrocitos a la lisis mediada por células que es característica en la hemoglobinuria paroxística noctura, un defecto adquirido que afecta a células hematopoyéticas con relación clonal. Actualmente se sabe que la hemoglobinuria paroxística noctura es causada por una mutación somática en las células hematopoyéticas tronco a nivel del gen PIG-A del cromosoma X. El producto del gen PIG-A actúa en la biosíntesis del anclaje glucofosfatidilinositol (GPI) de las proteínas de la membrana. Por lo tanto, la progenie celular de las clonas afectadas no tiene FAD ni CD59, las dos proteínas de anclaje-GPI que protegen a las células del ataque del complemento.
Algunos pacientes con glomerulonefritis membrano-proliferativa y la mayoría de los pacientes con lipodistrofia parcial con o sin glomerulonefritis, muestran un mecanismo poco frecuente de activación de la vía alterna del complemento mediada por anticuerpos. Estos individuos muestran un perfil del complemento sérico indicativo de activación de la vía alterna, es decir, niveles bajos de C3 y concentraciones normales de C1, C4 y C2. En el suero de estos pacientes también puede detectarse un anticuerpo de IgG, denominado factor nefrítico C3, que es específico para determinantes antigénicos expresados por la amplificación de la convertasa de C3. Una molécula de factor nefrítico C3 se une a una de C3b,Bb, creando una enzima trimolecular estable. Una vez estabilizada de ese modo, la convertasa de C3 amplificadora puede resistir la disociación de la subunidad Bb mediada por H y activar sin obstáculos la secuencia terminal del complemento.
Actividad biológica del sistema del complemento
Los fragmentos proteolíticos de los factores del complemento, sobre todo de C5a, desempeñan papeles importantes en la inflamación aguda. C5a, mediante la unión a receptores específicos, proporciona la señal para que los neutrófilos, monocitos y otros leucocitos migren hacia los sitios de activación del complemento. La unión de C5a aumenta el número de receptores sobre monocitos y neutrófilos que identifican otros factores, incluyendo C3b e iC3b. C5a y C3a estimulan a las células cebadas para que liberen histamina y otros moduladores de la inflamación, que ayudan al movimiento de neutrófilos a través del endotelio. C3e, un fragmento de C3b, incrementa el número de neutrófilos en la sangre periférica disponibles para ser reclutados a los sitios de activación del complemento, al promover la liberación de estas células a partir de la médula ósea y las reservas marginales.
La actividad de estos fragmentos del complemento suele ser benéfica cuando se relaciona con la defensa contra la infección microbiana. Sin embargo, estas acciones pueden ser dañinas cuando se asocian con activación excesiva del complemento en forma local o sistémica. Por ejemplo, ciertos tipos de membranas de diálisis activan la vía alterna y generan grandes cantidades de C5a, lo que provoca la formación de agregados leucocitarios que son atrapados de manera transitoria en el lecho capilar pulmonar. La producción excesiva de C5a y el atrapamiento de leucocitos también contribuye al daño endotelial pulmonar que ocurre en pacientes con el síndrome de insuficiencia respiratoria del adulto. Otro ejemplo se observa en pacientes con quemaduras, que muestran activación extensa del sistema del complemento por el tejido lesionado; los neutrófilos son activados en forma sistémica por la producción excesiva de C5a. Es probable que los efectos favorables de la supresión de C3 en modelos animales de infarto agudo del miocardio resulten de la ausencia en la producción de C5a y la menor infiltración del miocardio isquémico por neutrófilos.
Los receptores de C3b, C3dg e iC3b se denominan receptores del complemento 1, 2 y 3 (CR1, CR2 y CR3), respectivamente [ver tabla 2]. Estos receptores son importantes porque los fragmentos de C3 con los que interactúan se unen a los sitios blanco de la activación del complemento, como bacterias o complejos inmunes. Por ejemplo, CR1 y CR3 reforzan la capacidad de neutrófilos, eosinófilos y monocitos para fagocitar bacterias cubiertas con C3b e iC3b. La unión de C5a aumenta la concentración de CR1 y CR3. La mayor expresión de CR3 también promueve la adherencia de los neutrófilos a las células endoteliales, un paso fundamental en la localización de los neutrófilos en el sitio de la activación del complemento. El papel crucial del CR3 se demuestra en pacientes con deficiencia hereditaria de CR3, que sufren infecciones bacterianas repetidas, debido a que está deteriorada la capacidad de los neutrófilos para acumularse en el sitio de la infección.
|
||||||||||||||||||
|
CR1 también permite la unión de los eritrocitos a complejos inmunes circulantes que se unen a C3b, y con ello evitan la difusión de estos complejos a los tejidos. La unión de C3b a CR1 causa la formación de C3dg, que es transferido al CR2. Este último se asocia con el CD19, una proteína de membrana de las células B que disminuye el número de receptores antigénicos que deben estar ocupados para que ocurra la activación celular. La consecuencia de estas interacciones es una respuesta intensa de anticuerpos a dosis bajas de antígenos. Resulta interesante señalar que la molécula CR2 de los linfocitos B sirve como receptor para el virus de Epstein-Barr (VEB), modulando la infección y la inmortalización (el VEB se relaciona con el linfoma de Burkitt en Africa y con el carcinoma nasofaríngeo). El papel primario del CR2 en la patogenia de la infección por el virus de Epstein-Barr se demuestra por la observación de que las células L murinas que se han transferido con ADNc para el CR2 son susceptibles a la infección por VEB.
Dos de los efectos biológicos del complemento no incluyen receptores específicos de membrana: la citólisis de las células blanco por la introducción del complejo C5b-C9 en la membrana celular y el mantenimiento de la solubilidad de los complejos antígeno-anticuerpo. El primer mecanismo representa una vía por la cual el sistema del complemento contribuye a la defensa del huésped, mientras que el último resulta importante en la prevención de la enfermedad por complejos inmunes; ambos mecanismos requieren que se encuentre intacta la vía clásica de la activación del complemento.
En resumen, el sistema de complemento, activado ya sea por anticuerpos o por la vía alterna, entraña una serie de reacciones vitales para la defensa del organismo. Este grupo de reacciones produce inicialmente un aumento en la permeabilidad vascular, lo cual permite que una mayor cantidad de componentes del complemento y anticuerpos penetren al sitio de reacción y amplifiquen el proceso de activación. El componente activado C5a atraerá entonces células efectoras como neutrófilos y monocitos, y la unión de C3b al complejo antígeno-anticuerpo extraño facilitará aún más el proceso de fagocitosis. Algunos organismos y células son lisados directamente. Sin embargo, al igual que los sistemas de defensa humoral y celular, el sistema del complemento es como una hoja de dos filos: puede aumentar la resistencia del organismo a la infección, pero también puede producir daño tisular.
Calten HR, Rasen FS: Camplement deficiencies. Annu Rey
Immunal 10:809 ,1992
Fearon DT: The CD19-CR2-TAPA-1 complex, CD45 and signaling
by the antigen receptor of B lymphocytes. Curr Opin ImmunoI5:341, 1993
Fearon DT, Aheam JM: Complement receptor type 1 (C3b/C4b
receptor; CD35) and complement receptor type 2 (C3d/Epstein-Barr virus
receptor; CD21). CurrTop Microbiol ImmunoI 153:83, 1990
Figueroa JE, Densen P: Infectious diseases associated
with complement deficiencies. Clin Microbiol Rev4:359, 1991
Frank MM: The mechanism by which microorganisms avoid
complement attack. Curr Opin ImmunoI 4:14, 1992
Takeda J, Miyata T, Kawagoe K, et al: Deficiency of the
GPI anchor caused bya somatic mutationof the PIG-Agene in paroxysmal noctumal
hemoglobinuria. Cell 73:703, 1993
Tomlinson S: Complement defense mechanisms. Curr Opin
ImmunoI 5:83, 1993